Reconstructing genetic history of Siberian and
Northeastern European populations. 2017.1.7
2012년말에
출간한 "유라시아어의 기원과
한국어"에서부터 필자가
줄곧 강조한 것은 인도유럽어가 우리말과 같은 언어적 기반에서 기원했다는 것이었습니다. 영어가 우리말과 같은 토대 위에 있다는 주장이 되므로,
지금까지 교육 받은 것과는 너무도 달라, 일반 상식으로는 도저히 이해할 수 없는 내용이었죠. 현대 영어는 우리가 익히 알듯이 우리말과 너무도
다릅니다. 그러나 이를 소급해 올라 가면, 결국 '원인도유럽어(=PIE,Proto Indo Euroup Language)'를 만나게 되고, 이렇게 재 구성된PIE
언어는 우리말의 그것과 점점 가까워지게 된다는 겁니다.
기원 전후 만하여도 이미
상당 부분 어지러워진 희랍어와 라틴어만 보아서는 이를 확증할 수 없지만, 그보다 오래전에 고정된 산스크리트어를 본다면, 이의 연관 고리에 대한
확신이 생깁니다. 그리고 인도유럽어는 지난 150년간 엄청나게 이런 언어 간의 관계를 연구하여 매우 강력한 데이터 베이스가 이미 구축되어
있습니다. 인터넷에 etimology를 치면, 관련 사이트들이 쭉 나옵니다. 따라서 이제는 이런 데이터들이 있기 때문에, 거짓말을 할래야 할
수가 없습니다. 그런 데이터가 널려 있는데도 불구하고, 이런 사실들에 대한 자각이 전연 없다는 게 우리들의 이상한
현실입니다.
아무튼 이러한 언어
분석에 의하면, 오스트로네시안 언어가 우리말의 기층이고, 인도 유럽어는
우리말과 같은 토대 위에 성립된 언어라는 것이 필자가 주장하는 골자입니다. 말하자면 1층은 오스트로네시안
언어이고, 2층은 인도유럽어라면, 우리말은 1층과 2층을 연결하는 복도 계단이 되는 겁니다.
이런 필자의 듣보잡
주장은, 2015년부터 고인골 유전자 분석이 가능해지기 시작하면서, 그 비밀이 하나씩 풀리게 됩니다. 즉 이상하게도 이런 고인골 유전자 분석
결과는 발표하는 것마다 필자가 기존에 주장하던 언어학적인 주장을 하나씩 확인해 주고 있기 때문입니다. 유럽인들이 볼가강 근처
얌나야 문화에서 기원했다는 것이 2015년 2월 밝혀지면서, 현재 유럽 제 언어들과 PIE 간에 불분명하던 그 하나의 고리가 확인되면서, 그간
수십년간 고고학계와 언어학계의 화두였던 '아나톨리아(Anatolia)
기원설'과 '흑해 북단 기원설(=Pontic
Steppe)' 논쟁은 단칼에 '흑해 북단
기원설'로 정리되고 맙니다.
그 이후로 좀 더 조사를
해보니, 이 흑해 북단인들인 얌나야인들은 예상과는 전혀 다르게 바이칼의 말타부렛(Malta-Buret)에서 유래했다는 것이 추가로 밝혀
집니다. 이리하여 유럽인들은 기원이 고-시베리아란 것이 밝혀집니다. 이는 기존의 통념과는 완전히 배치되는 내용이죠. 그러나
그럼 과연 "이
'고대 북유라시아인(=ANE:Ancient North Eurasian)'들은
어디서부터 기원하는 걸까"라는 마지막 연결 고리는
여전히 미완인 채 남아
있었습니다.
아무튼 1차
'얌나야(=Yamnaya)' 고리와 2차
'알타이/바이칼' 고리가 밝혀 질 때 만하여도,
유럽인들도 전연 예상치 못한 이런 고인골 유전자 분석 결과를 두고 온갖 해석이 난무했습니다. 일반 고정 관념에 따르면, 메소포타미아에서 당연히
기원을 했어야 하죠. 이런 고인골 유전자 분석 결과들은, 기존의 서양 사회의 믿음과 전면 배치되는 내용인지라, 처음에는 이를 받아 드리기 어려워
했지만, 이제는 거의 교통정리가 되어, 이제는 이를 과학적 사실로 받아 드리는 듯 합니다. 없던 역사까지 만들어 역사 왜곡을 하는 중국이나
일본과는 달리, 역시 사실을 사실로 받아 드리는 서양의 열린 사회의 모습을 보는 것 같아 보기가 좋습니다.
아무튼 필자는 이런 최근
유전자 분석 이전에, 고대 언어 연구에서 나온 결과가 그러했기 때문에, 당연히 언어에서 보여 주는 방식 그대로, 유전자도 그런 연관 고리를
가져야 한다고 일관되게 주장을 했었던 거죠. 그러나 사람들에게는 필자의 이런 주장이 너무나 일반 상식과는 너무나 동 떨어진 얘기인 지라, 다들
무슨 그런 개소리냐고 여겼겠지만 말이죠.
하지만, 이번에 발표된
러시아 논문에서는 2.4만년전의 바이칼 '말타
부렛(Malta-Buret)'과 1.7만년전의 알타이 지역의 '아폰토바 고라(Afontova-Gora)' 유적에서 발굴된
고인골과 현재 유럽인들, 그리고 시베리아 소수 원주민의 유전자를 비교하여, 남아 있던
마지막 고리로, 이들이 동 시베리아에서 기원했고, 나아가 이들이 동북
아시아가 유전적 조상이라는 것을 확인해주고 있습니다. 물론 이 논문에는 이런 동북아 기원설을 명시적으로는 언급하지 않고 자기들 입맛에 맞게 다르게 비틀어서 말하고
있지만, 이 논문을 보면,
바로 이런 내용이 핵심이라는 것을 알 수 있습니다. Yamnaya건도
처음에는 못 믿어워 비틀어 말했지만, 결국은 많은 물증이 후속적으로 분석됨에 따라 결국은 '말타 부렛(Malta-Buret)'과 이 현대
유럽인들의 직접적인 조상이라는 것을 인정하고 싶지 않았지만, 결국 인정하게 된 겁니다. 아무튼 이런 고인골 유전자 분석 결과물들은 필자가 언어
연구를 통해 이미 5년전에 제시한 '유라시아
패러다임'이라는 거대한 인류 이동 프레임을 재확인시켜 주는 고인골
유전자 분석 결과입니다. 사필귀정인
거죠.
3만년전에 오세아틱
유전자에 기초하여 동남아시아에서 출발하여 유럽으로 이동하여 초기 유럽인들을 형성하게 됩니다. 이들 루트는 필자가 주장하는 경로에 따르면,
동남아에서 출발하여, 중국 해안가를 따라 북상하다가, 한반도 지역에서 툰드라 지역에 진입하게 됩니다.
툰드라로 진입하면서
채집 생활에서 수렵 생활로 생활 양식이 완전히 바뀌게 됩니다. 물론 이런 환경 변화는 격언어라고 지칭 되어지는 목적어 우선 언어의 탄생을
동반합니다. 이들은 유라시아 대륙 동쪽에서부터 유라시아 대륙을 뒤덮고 있던, 거대 빙하 자락에 펼쳐진 광활한 툰드라 벨트를 따라 서유럽에까지
이르게 됩니다. 이들이 '고대 북유라시안(ANE:Ancient Nprth
Eurasian)'을 형성하게 됩니다. 이런
툰드라 지역의 진입은 5만년전에 이루어졌고, 이후 이 지역에서 4만년간 유전적으로 분기하고 또 성장해 나갑니다.필자는 이 시점에서
유라시아 대륙의 '툰드라(=Tundra) 벨트'가 유전자와 언어
분기의
요람(=Basin)을 이루었다고 주장합니다. 3만년 이전에
이렇게 유럽에 도달한 이들을 고고학에서는 유럽의 Augrinac 구석기인이라고
지칭합니다.이들은 3만년 이후
2만년전까지는 Gravettan 구석기인들로 대치되었다가, 최대 빙하기 기간을
거치면서, 이 gravettan인들은
거의 망하고, 툰드라 벨트를 의지하면서 추위에 잘 적응한 Augrinac인들이 피한지에서 살아 남아 최대 빙하기 이후에 컴백하면서,
이후
'유럽 토착 수렵채집인(=WHG, West Hunter Gatherer)'들을 형성합니다. 아무튼 이 과정에서, 일부 외부 유전자 유입이 있었지만, 대체적으로 보면, 초기 진입한 이들이 주축이 되어 유럽과 시베리아에서 자체 분기하고 성장합니다.
피부색은 당연히 검은 색입니다.
이 중에 바이칼과
알타이 지역에서 새로이 현대 유럽인들 유전자를 가진 남성의 조상이 출현합니다. 이들은 툰드라 벨트를 의지하여 성장하다가, 빙하기가 물러 나면서
지금과 달리 살기 좋아진 시베리아를 배경으로 지속적으로 성장하게 됩니다. 이들은 알타이-우랄 지역에서 성장하고, 이후 이들이 청동기 때 이르면,
볼가강 지역에서 어떤 이유인지는 몰라도 청동기, 수레, 말과 함께 대거 이동을 합니다.
이들이 가지고 간
문화가 토기로는 시베리아 빗살무늬 토기에 기초한 CordedWare 토기, 매장 방식으로 '쿠르간(=Kurgan)의 무덤', 그리고 언어로는 ' PIE(=Proto Indo Euroupean)'입니다. 이렇게
이동하여 유럽으로 진입하여 현재의 유럽인들의 유전적 구성을 이때 형성하게 됩니다. 이때 아마도 '원인도유럽어(=PIE)'라는 언어도 같이 동반한 듯 싶습니다. 유럽의
제 언어는 수많은 변형을 거쳤지만, 이 '원인도유럽어(=PIE)'에서 기원합니다. 인도유럽어의 간에
보이는 언어의 일관성은 바로 이런 이유로 설명이 가능합니다. 즉 빙하기 때 동으로 한반도에서부터 서로는 이베리아 반도에 이르는, 유라시아 대륙을
동서로 가로지르는 거대한 툰드라 벨트 지역이
그 토대가 되었다는 것이 필자의 주장입니다. 이것이 바로 필자가 주장하는 '유라시아 패러다임'의 골자입니다.
물론 현재의
영어나, 유럽의 제 언어는 우리말과 매우 이질적으로 다릅니다. 그러나 이는 청동기 이후의 언어로 인류가 만든 온전치 못한 문자와 잘못된 언어
규칙을 잘못 적용하여 언어가 망가진 것으로 필자는 파악하고 있습니다. 인도 유럽어는 역사적으로 발전한 것이 아니라, 오히려 인위적으로 잘못된
것들의 오염으로 타락한 것이라는 거죠. 물론 현재의 학교 교육에서 배운 바로 보면, 이런 주장을 믿을 수 없겠지만, 이는 필자가 주장하는 핵심
내용 중에 하나입니다. 그러나 이렇게 유럽의 제 언어가 인위적으로 언어적 타락을 하기 이전의 고대 언어 형태, 예를들자면 기원전 언어 형태를
고스란히 간직하고 있는 산스크리트어, 희랍어, 고전 라틴어를 살펴 보면, 이들이 서로 매우 유사한 언어 형태를 지니고 있음을 보게
됩니다.
그리고 이들 고대
언어가, 지금껏 학교에서 배운 내용과는 전연 다르게, 이상하게도 우리말과 그 맥락이
닿아 있다는 것을 필자는 발견하고, 왜 이런
언어적인 일치성을 지닌 지를 그 메커니즘을
밝힌 것이 2012년 발간한 제 1권
"유라시아어의 기원과 한국어", 제2권
"한국어로 재 정립한 범어 문법", 제3권 "범어 금강경 직독과 문법 해설"입니다. 이는 약
1800페이지에 이르는 분량의 저작입니다.
후속적으로 인도 유럽어와 우리말이 어떻게 연계가 되느냐 하는
점은 낱낱이 밝힐 수도 있겠지만, 당분간은 여백으로 남겨 둘 예정입니다. 이미 고기 잡는 법을 제시했기 때문에, 고기를 많이 잡아 갖다 주는
것이 아니라, 스스로 고기를 잡도록, 즉 스스로 알아 내도록 하는 겁니다. 다 아르켜 주면, 스스로 발견하는 재미가 없기 때문입니다. 지금은
이런 사소한 것에 시간을 소비할 여력이 없습니다. 조만간 이보다 매우
강력한 것을 제시할 예정으로, 이는 새로운 언어 시대를 열 것입니다.
그러나 이런 결론이
나왔지만, 왜 이런 동서양 언어 간에 이런 언어 현상을 갖는가가 필자에게는 여전히 매우 큰 의문으로 다가 왔습니다. 지리적으로 5천km나 떨어진 곳에서 이런 언어의
일관성을 보인다는 것은, 필자 스스로도 도저히 납득이 되질 않는 겁니다. 따라서 이런 의문을 답하기 위해 보다 더 정확히 대답해 줄 수 있다고
여겨지는 유전자 분석에 관심을 갖는 계기가 되었습니다.
그런데 최근에 2015년부터 이런 고인골 유전 분석이
드디어 가능해지면서, 속속 인류의 여정이 과학적으로 밝혀지기 시작합니다. 그런데 여기서 정작 중요한 것은 필자가 언어분석을 통해 이미 예측한
결과를 이 고인골 유전자 분석 결과가 거꾸로 하나씩 확인시켜 주고 있다는 점입니다. 필자가 무슨 점쟁이도 아니지만, 웬일인지 모르지만, 필자가
언어분석으로 예측한 것이 그대로 고인골 분석 결과로 밝혀지고 있다는 게 필자 스스로도 믿을 수 없을만치 그저 놀라울
뿐입니다.
필자의 가설은
대부분은 적중하여 2015년부터 유전자 분석 결과가 확인해 주었지만, 필자의 주장 중에서 아직 밝혀지지 않는 게, "툰드라 가설"입니다. 이 가설의 중심에는 동북 아시아가 툰드라 지역의
유전적 확산 시초가 되어야 한다는 점이 핵심 포인트입니다.
이에 관해 답해 줄
수 있는 내용이 바로 이 논문입니다. 물론 이 논문을 쓴 사람은 이런 취지를 부정하고 싶겠지요. 이 논문의 결론
부분의 그려 논 그림을 보면, 논문
저자들의 이런 의도가 그대로
보입니다만, 사실 이
논문이 말해 주는 것은 바로 '동북아 기원설'이 그
핵심입니다.
사실 필자는 이런 유전적
고리가 언어 분석처럼 유라시아 대륙을 관통하여 반드시 있다고 여겼지만, 이런 연관성을
입증할 시베리아 소수 원주민들의 유전자 정보가 턱없이 부족하였기 때문에, 이를 확증할 수 없어 늘 목말라 했습니다. 그러나 러시아가 드디어 이
논문을 통해 그 전모를 제시하였습니다. 이로써 필자가 주장한
언어학적 주장이 유전자 분석 결과와도 일치한다는 것을 이것으로 최종 확인하면서, 그 유라시아 언어의 퍼즐 조각이 유전자와 더불어 모두 맞춰지게
된 것으로 보입니다.
아무튼 이것이 의미하는
것은 우리말이 인도 유럽어의 언어적 원형에 해당한다는 얘기입니다. 물론 어떻게 하여 그리되었는지 몰라도, 우리말에는 인도 유럽어의 원형이 가장
많이 남아 있다는 얘기입니다. 필자가 보기에는 아마도 한반도라는 독특한 지리적 고립성 때문인 것 같습니다. 정작 유럽인들은 그들의 언어를 아무리 드려다 보아도 이해할 수가 없습니다. 한국어를 통하지
않고는 그들 언어의 기원을 도저히 알아 낼 수 없다는 얘기입니다. 우리 스스로도 지금껏 몰랐지만, 한국어에는 이런 게 들어
있습니다.
한편 이런 고리타분한
옛날 얘기, 이것이 왜 현대의 우리에게도 중요하냐 하면, 우리가 누리는 문명이 바로 이런 언어적 토대 위에 서 있기 때문입니다. 현대 기술
문명을 이룩한 언어는 누가 뭐래도 '인도유럽어' 입니다. 지금 우리가 향유하는 현대문명은 바로 인도
유럽어의 언어 패러다임 위에 구축돼 있다고 해도 과언이 아닙니다. 덕분에
현대에 일반인들인 우리는 조선시대 때 왕들보다 더 나은 수준의 물질 문명의 혜택을 누릴 수 있게 되었습니다. 이에 대한 고마움은 잊어서는 아니
됩니다.
이러한 물적 토대의
대대적이 확충에도 불구하고, 오히려 반대로 출산율의 저하, 결혼의 포기, 취업 포기 등과 같은 사회적 현상이 만연하고, 사람들이 행복하질 않는
겁니다. 물적 토대는 전 시대에 비해 엄청나게 풍요해졌지만, 이상하게도 이러한 물질적 풍요와는 반대로, 오히려 생활에 여유는 더욱더 없어지고
삶의 쪼들림은 점점 심해져 간다는 겁니다. 말하자면, 대공황 때 바로 옆에 식량을 쌓아 놓고도, 굶어 죽어 나간 경우와 같은 어처구니 없는
현상이 생기는 겁니다. 이런 게 문제라하여 공산주의도 해 봤지만, 더 문제가 많다는 것만 확인해 준 겁니다. 이미 자본주의라는 전제 위에 안티
테제로 출발한 것은, 지킬 박사와 하이드처럼, 한 몸체의 양쪽 얼굴일 뿐입니다. 즉 공산주의는 자본주의와 사실은 같은 거라 보시면 됩니다.
동전의 양면과 같은 것으로, 이는 사실은 같은 패러다임이라는 거죠.
아무튼 이런 사회현상을
가리켜 '헬조선'이라 칭하지만, 이는 비단 우리만의 문제가 아니라, 여러나라가 동시에 겪고 있는 공통적인 현상입니다. 아무리 천재적인 경제학자가
와도 이런 문제 해결에는 전연 도움이 안 될 뿐만 아니라, 오히려 오진을 해 처방해서 아예 망쳐 놓기 일쑤입니다.
그러나 우리는, 이런
여러 현상이 매우 복잡 다단하지만, 바로 다름 아닌 우리 자신이 그렇게 만들었다는 것 만은 명확히 알 수 있습니다. 이 배경에는 바로 언어의
그림자가 드리워져 있기 때문입니다. 우리는 '우리가 하는 사고'와 그리고 만들어 내는 '사회 형태'와 '행위의 모든 행태'는 언어 공간 위에
구축되어 있습니다.
우리는 이를 사회적으로
'공유'를 합니다. 이는 게임의 세계처럼 실재하는 것은 아니지만, 게임 안에서는 게임이 실재가 되는 것처럼, 우리에게는 이러한 것은 실체로
작용합니다. 예컨대 빚을 지면 반드시 갚아야 한다든가 하는 것 등이 성립되는 거죠. 이런 모든 것들은 '두 가지 상태로 기술하는 것'으로부터
구성돼 있습니다.
이 '두 가지 상태로
기술하는 것'의 총아는 바로 다름 아닌 '컴퓨터'입니다. 영어는 역사적으로 보면, 이 두 가지 상태로 언어가 최적화 된 겁니다. 덕분에 2가지
상태로 기술되는 세계에서는 매우 유럽문명이 성공적일 수가 있었습니다. 그러나 이 두 가지 상태란 것도 실재하는 것이 아니라, 우리가 그려 낸
거라는 겁니다. 예컨대, 바둑의 세계에서 흑백은 바둑판에서만 유효한 것일 뿐인 거와 같습니다. 바둑 세계 안에서만 유효한 거와
같습니다. 그러나 이 두 가지 상태는 무한한 바둑 세계를 만들어 냅니다. 수 많은 좋은 수와 나쁜 수를 만들어 냅니다.
서양에서 과학이 발달한
이유는 두 가지 상태로 언어를 운용하였기 때문에 그렇습니다. 문제는 여기까지는 괜찮은데, 마치 바둑에서 대마가 얽히듯이, 사회가 엉키는 게
문제가 됩니다.
즉 필자는 이를
'Dead-Lock'이라 칭합니다. 물질의 풍요에 불구하고,
'삼포' 세대가 되어야 하는 것은 바로 이런 'Dead-Lock' 현상이기 때문입니다. 역사적으로 자본주의가 문제가 있다
하여 공산주의를 하여 문제가 해결된 적은 한번도 없습니다. 공산주의와 자본주의는 같은 패러다임 위에 서 있는 동전의 양면과 같은 것입니다. 이는
빛과 그림자처럼 서로를 의지해 존재합니다. 다시 말하면, 문제를 해결한다고 그 반대를 시행하여 문제가 해결되는 게 아니라는
얘기입니다. 지금 수 많은 천재적인
경제학자가 이런 문제를 풀려고 하지만, 불행히도 이런 근본적인 문제를 통찰하지 못하기 때문에, 영원히 풀 수 없는 문제가
됩니다.
예를들면 최저임금 문제를
들 수가 있겠습니다. 최저임금을 높이자는 것은 좌파고 내리자는 것은 우파입니다. 그러나 이를 높이거나 내리는 것으로는 문제 해결이 안 된다는 것
만은 자명합니다. 최저 임금을 높이자는 방향성을 맞지만, 그러나 그렇게 하면 결국은 무인 시스템 도입의 기폭제로 밖에는 작용하지 않는다는
겁니다. 즉 결론적으로는 무인화를 급속히 앞 당기는 결과가 될 뿐이라는 겁니다. 즉 사람 중심이 아닌 기계 중심 세상으로 이끄는 출발점이 될
뿐이다.
이제는 노동은 과거처럼
노역으로 하던 시대는 점점 저물어 간다. 이런 노역은 기계가 대신 할 것이다. 따라서 사람들끼리 어울려 살아 가는 메카니즘을 만들어 내야 한다.
최저임금을 올리거나 내리는 것과 같은 동전의 양면을 붙들고 영원히 해결될 수 없는 싸움을 싸울게 아니라, 실은 어울려 살아 갈 수 있는
메카니즘을 구현해야 한다. 일반 사람들의 언어 구조는 이분법인지라, 그에 맞는 말을 해 줘야 인기를 얻게 된다. 올리거나
내리거냐이다.
그러나 그것은 불행히도
그건 해결책이 아니다. 달콤한 프로파겐다치고 성취된 것은 아무것도 없다. 실은 전연 예상치 못한 곳에서부터 실마리가 풀렸다. 중국을 보면 그
실마리가 전혀 예상치 못한 곳에서부터 풀어지고 있다. 대약진 운동과 홍위병이 성취한 것은 아무것도 없다. 오히려 수백만명의 아사자와 수천만명의
맞아 죽은 사람만 만들었을 뿐이다. 지금은 그래도 중국은 먹고 사는 문제에 있어서는 조금 희망은 있어 보인다. 중국 지도부가 요즘하는 얘기를
보면, 어디서 세뇌를 받았는지는 몰라도 상당한 수준이다.
그런데 문제는 올리거나
내리거나를 주장해야 하는 정치 패러다임과 다른 딴소리하는 정치인은 그 정치판에서는 반드시 도태가 된다는 점이다. 왜냐하면 정치판 구조에 맞지
않기 때문이다. 그렇기 때문에 정치에 뭤을 기대하는 것 자체가 실은 연목구어가 된다. 이런 판 안에서는 태생적으로 기대할 것이
없다.
따라서 우리는 이제
패러다임의 쉬프트를 통해, 문제 해결에 나서야 합니다. 새로운 사회 Space를 구현하는 것이 그 해결책이다. 패러다임을 바꾸고, 새로운
메커니즘을 만들어 냄으로써 구현이 된다. 이는 좌파의 패러다임이나 우파의 패러다임에 속한 것이 아니다. 촛불은 원래 국민적 열망인데, 불행하게도
좌파 프레임으로 추락하여 패러다임 쉬프트의 원동력을 상실하고 말았다. 안타까운 일이다. 태극기와 촛불은 동전의 양면으로 프레임에 갇힌 것이다.
그러나 둘다 실은 같은 열망에서부터 출발했을 것이다. 서로가 서로를 마녀 사냥을 하여 자기의 정당성을 확보할 게 아니라, 서로가 같은 열망을
공유하였다는 측면에서 존중하면서 서로의 목소리를 내야 할 것이다.
아무튼 언어는 바로 이런
패러다임이 처음 발현되는 곳입니다. 자칫 잘못하면, 개인적으로도 이런 언어의 패러다임에 낚여, 평생을 언어의 노예로 살면서도 그런 줄을 모르고
살고 있다는 것도 우리는 이제 좀 알아야 합니다. 이런 게 실은 우리의 현실이라는 자각을 할 필요가 있다는 거죠.
개인적 뿐만이 아니라,
사회적으로는 또한 사회 시스템의 노예로 살아 갑니다. 이렇게 생성된 시스템은
'천국(Heaven)'이 아니라, '지옥(Hell)'로 작동될 수 있습니다. 헤븐'이던, '헬' 이던 간에,
중요한 것은 우리가 그렇게 만들었다는 게 중요합니다. 이렇게 되면, 물질의
풍요에 관계없이, 우리는 그런 관계를 형성하여, 아무도 시키지 않았지만, 스스로 그렇게 '헬'을 만들게
됩니다.
간단한 예를 보면, 학교 사회를 보면 됩니다. 현재 학교는 한 마디로 요약하면 점점 '정글화'가
되어 갑니다. 또한 우리는 우리도 모르게 GMO를 많이 먹어서 그런지는 몰라도 자폐증이나 ADHD 비율도 일반인들이 상상 이상으로 매우
높습니다. 곳에 따라서는 50%에 육박하는 학교도 있답니다. ADHD는 꼭 나쁜 것 만은 아닙니다만, 아무튼 그렇다는 거죠. 학교는 사회의
반영물이기 때문에, 학교 문제는 궁극적으로는 사회문제입니다. 학교 폭력을 비롯해, 많은 문제가 학생들 사회에 있습니다. 이런 환경을 겪은
사람들에게는, 부차적으로 이런 분위기에서는 애를 낳고 싶은 생각이 점점 들지 않겠지요. 이렇게 되니, 선생 해 먹기도 힘들어, 특히 중학교의
경우는 학폭 등으로 하루 하루를 조마조마 하게 보내고 있고, 여건만 된다면 선생을 아예 관 둘려고 한다고 합니다.
언어 분석을 해 보면,
원래 오리지널 '원인도유럽어(=PIE)'는 '두 가지
상태에 고착화'가 되지 않아 매우 유연했었습니다. 인류에게 있어, 붓다, 노자, 공자, 예수와 같은 대 성인들이 출현한 배경에는 당시의 언어들이
덜 오염된 탓도 있습니다. 같은 인도 땅에서도 AD7~8세기에 상키아 철학 같은 것이 나온 배경에는 변형된 언어 문제가 있기 때문입니다. 언어가
꼬였기 때문에, 이젠 꼬인 대다가 억지로 맞추어 얘기를 해줘야 그제서야 감동을 하게 됩니다. 이게 바로 샹키아 철학입니다. 그 옛날처럼 바른
얘기를 해 주어도, 이제는 공명이 일어나지 않아 대 성인이 출현하더라도 할 일이 없게 됩니다. 얼핏 들으면 매우 웃기는 얘기이겠지만, 이게
현실이라는 것을 우리는 분명히 알 필요가 있습니다 .
이처럼 지금 우리는 우리
스스로가 베베 꼬여 있습니다. 시스템도 베베 꼬여, 점점 Dead-Lock이 되어 가고 있습니다. 언어가 유연하다는
것은 새로운 패러다임의 세상 창출이
보다 쉽게 될 수 있는 좋은 기반이 될 수 있습니다. 한국어는 PIE 보다도 더 유연합니다.
따라서 필자는 한국어
사용자 사회에게 많은 기대를 하고 있습니다. 이제 우리는 지난 시대와는 달리, 아이디어만 있다면 바로 구현해 볼 수 있는 기술적으로 가능한
시대에 살고 있습니다. 이러한 얘기를 하는 이유는, 이러한 통찰을 가지고 일을 한다면, 지금 시스템의 패러다임의 관성을 넘어 새로운 패러다임의
세상을 열 수 있기 때문입니다. 문제 안에서는 문제가 결코 해결되지 않습니다. 오로지 패러다임 쉬프트 만이 가능성이 있습니다. 아무 생각이
없으면, 그냥 이전의 관성대로 그냥 흘러 갈 뿐입니다.
구글이나 아마존,
페이스북 등등은 이런 패러다임 쉬프트가 아니라, 어떻하면 돈을 더 많이 긁어 모을까에만 관심이 있기 때문에 이런 것들을 기대하기가 어렵습니다.
이런 회사들은 패러다임 쉬프트가
아니라 기존 패러다임을 확대
재생산하기 때문에 별로 가망이 없습니다.
이제 '인공지능'이라는
거대한 21세기형 선악과가 우리 앞에 던져져 있습니다. 이는 삼킬 수도 뱉을 수도 없는 것입니다. 이는 우리에게 '헤븐'을 갖다 줄 수도,
'헬'을 갖다 줄 수도 있습니다만, 지금과 같은 패러다임 진행 대로라면, 당연히 '헬'이 올 것으로 예상이 됩니다. 그 '헬'은 단순한 '헬'이
아니라, AI의 등장과 더불어 인류에게 종말을 가져다 줄 것입니다.
여기서 몸을 돌려,
패러다임의 전환을 한다면, 오히려 이런 21세기 선악과는 생명나무와 더불어 천국을 이루는 중요한 메카니즘이 될 것입니다. 알고 보면 매우 간단한
거지만, 그러나 이를 극복할 객관적 확률은 그리 높아 보이지 않습니다. 그러나 그렇다는 것 만은 우리는 반드시 알아야 할 필요가 있는데, 통찰이
있다면, 언젠가는 헤어날 길이라도 있겠지만, 만약 통찰마저 없다면, 영영 이런 질곡에서 헤어날 가망이 없기 때문입니다.
필자는 이런 측면에서
한국 사회가 미래 인류 시대에 하나의 가능성을 가진 사회라는 점을, 그리고 그것이 어쩌면 인류의 마지막 희망일지도 모른다는 것을 말해 주고
싶습니다.
(사족: 2017년
1월27일 발표된 논문이었는데, 이 발표를 모르고 있다가 2017/09/20일에야 관련 자료를 검색하다가 뒤 늦게 알아 내었습니다. 오타가
많아도 내용이 중요하기 때문에 우선 올리고 봅니다.)
추상
시베리아와 러시아 북서부에는 문화적으로나 언어 적으로
다양한 토종 민족이 거주하고 있지만이 지역의 사람들의 유전 적 변이와 역사는 크게 특징 지어지지 않습니다. 우리는 그 지역의 14 개 토착민에 속하는 28 명의
개인으로부터 깊은 전체 유전체 염기 서열 데이터 (~ 38x)를 제시한다. 우리는이 데이터 세트를 현대의 32 개 인간 고대 게놈과
32 개의 추가 인간 게놈으로 조합하여 북부 유라시아 토착민의 유전 적 역사를 재구성했습니다. 우리는 시베리아와 동아시아의 인구가 조상의 38 %와
현대의 자손이없는 것으로 여겨지 던 45,000 세의 Ust'-Ishim 개체를 공유하고 있음을 발견했습니다. 서쪽 시베리아 인은 24,000 년 된 시베리아 말타 소년
MA-1이 대표하는 고대 북 유라시아 인에게 그들의 조상의 57 %를 추적합니다. 동부 시베리아 인구는 1 만 ~ 1 천년 전에 다른
동아시아 인구와 분리 된 독특한 하위 계보를 형성했습니다. 또한 우리는 사마라, 카렐 리야, 헝가리 및 스웨덴의
시베리아 인과 동유럽 사냥꾼 수집가 (8000-6600 년전) 사이의 혼합물을 밝혀 냈습니다. Yamnaya 사람들 (5300-4700 년
전); 현대의 북동부
유럽인들. 우리의 결과는 시베리아와
북동부 유럽 인구의 유전 적 역사와 시베리아에서 유럽으로의 고대 유전자 흐름에 대한 새로운 통찰력을 제공합니다. 현대의 북동부 유럽인들. 우리의 결과는 시베리아와 북동부 유럽 인구의 유전 적
역사와 시베리아에서 유럽으로의 고대 유전자 흐름에 대한 새로운 통찰력을 제공합니다. 현대의 북동부 유럽인들. 우리의 결과는 시베리아와 북동부 유럽 인구의 유전 적
역사와 시베리아에서 유럽으로의 고대 유전자 흐름에 대한 새로운 통찰력을 제공합니다.
논문요약:
시베리아와
북서부 러시아에는 문화적으로나 언어적으로 다양한 40개 이상의 원주민들로 토착 소수민족들이 거주하고 있지만, 지금 껏 이 지역의 사람들의 유전적
변이와 그 변천 역사는 연구되지 않고 있습니다. 우리는 그 지역의 14 개 토착민에 속하는 28명으로부터
분석한 전체 유전체 염기 시퀀싱 데이터(~38x)를 제시합니다.
우리는 이
데이터와 현대의 32 개 인간 고대 게놈과 32 개의 현대인 게놈과 46개의 고인골 게놈을 결합하여, 북부 유라시아 토착민의 유전적 역사를
재구성했습니다. 우리는 현대의 후손이 없는 4.7만년전의
Ust'-Ishim 시베리아
고인골과 동아시아 인이 38%의 조상 유전자를 공유하고 있음을
발견했습니다. 서 시베리아 인은
2.4만년전 바이칼 말타 부렛 고인골인 말타(MA-1)로 대표되는 ANE(=고대 북 유라시아 인)에게 그들의 조상의 57%를 공유함을 알아
내었습니다. 현재 동-시베리아인들은
적어도 1만년 이전에 다른 동아시아 인구와 분기한 유전적 하위 계보를 형성합니다. 또한 우리는 Samara, Karelia, 헝가리 및
스웨덴의 시베리아인들에서 분기한 동유럽 수렵채집인 (8천-6.6천년전) 사이의 혼혈을 밝혀 냈습니다. 또한 흑해 북단의 Yamnaya 사람들 (BC3.3천-BC2.7천
년전)과 현대의 북동부 유럽인들도
혼혈이 발생한 것을 밝혀 내었습니다.
우리의 결과는
시베리아와 북동부 유럽 인구의 유전자 형성 역사와 시베리아에서 유럽으로의 고대 유전자 흐름에 대한
새로운 통찰력을 제공합니다.
추상
시베리아와 러시아 북서부에는 문화적으로나 언어 적으로
다양한 토종 민족이 거주하고 있지만이 지역의 사람들의 유전 적 변이와 역사는 크게 특징 지어지지 않습니다. 우리는 그 지역의 14 개 토착민에 속하는 28 명의
개인으로부터 깊은 전체 유전체 염기 서열 데이터 (~ 38x)를 제시한다. 우리는이 데이터 세트를 현대의 32 개 인간 고대 게놈과
32 개의 추가 인간 게놈으로 조합하여 북부 유라시아 토착민의 유전 적 역사를 재구성했습니다. 우리는 시베리아와 동아시아의 인구가 조상의 38 %와
현대의 자손이없는 것으로 여겨지 던 45,000 세의 Ust'-Ishim 개체를 공유하고 있음을 발견했습니다. 서쪽 시베리아 인은 24,000 년 된 시베리아 말타 소년
MA-1이 대표하는 고대 북 유라시아 인에게 그들의 조상의 57 %를 추적합니다. 동부 시베리아 인구는 1 만 ~ 1 천년 전에 다른
동아시아 인구와 분리 된 독특한 하위 계보를 형성했습니다. 또한 우리는 사마라, 카렐 리야, 헝가리 및 스웨덴의
시베리아 인과 동유럽 사냥꾼 수집가 (8000-6600 년전) 사이의 혼합물을 밝혀 냈습니다. Yamnaya 사람들 (5300-4700 년
전); 현대의 북동부
유럽인들. 우리의 결과는 시베리아와
북동부 유럽 인구의 유전 적 역사와 시베리아에서 유럽으로의 고대 유전자 흐름에 대한 새로운 통찰력을 제공합니다. 현대의 북동부 유럽인들. 우리의 결과는 시베리아와 북동부 유럽 인구의 유전 적
역사와 시베리아에서 유럽으로의 고대 유전자 흐름에 대한 새로운 통찰력을 제공합니다. 현대의 북동부 유럽인들. 우리의 결과는 시베리아와 북동부 유럽 인구의 유전 적
역사와 시베리아에서 유럽으로의 고대 유전자 흐름에 대한 새로운 통찰력을 제공합니다.
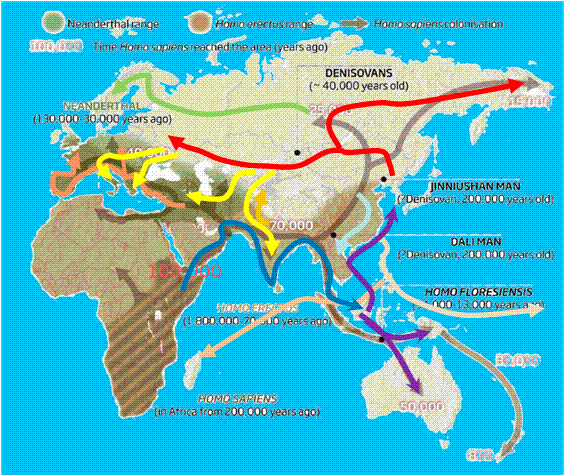
시베리아는 수십만 년 동안 원시
고대인들이 살았고, 고고학적 유적지 중 일부는 26만년 이상이나 된 것도 있습니다( Waters et
al. 1997 ). 네안데르탈 인은 유럽과 시베리아에 4만년 전
( Hublin
2009 )까지 살았고, 현생인류가 이 지역으로 확산해 들어 왔습니다
( Hublin
2012 ). 시베리아에 있는 현생인류의 거주 흔적은 서쪽 시베리아에
있는 Ust'-Ishim 유적지 근처에서 최근에 발견된 뼈로 미루어 알 수 있고, 또 이 뼈를 분석하여( Fu 등
2014 ) 네안데르탈인과의
혼혈이 50-60
kya사이에 있었음도
밝혀 집니다. ( Fu et al.
2014). 시베리아에 있는 다른 고대 인류 유적지들에서도 고대
고-유라시아인=(ANEs) ( Lazaridis 외
2014 )의 고대 DNA를 검출했으며, 구석기 시대 2.4만년전의 시베리아 Mal'ta 소년
MA-1 ( Raghavan 외
2014 )과 1.7만년전
아폰토바 고라-2 ( Raghavan et
al. 2014 ) 유적의 인골을 분석하여. 이 빙하기 툰드라 ANE 사람들로부터 서-시베리아인,
유럽인 및 아메리카 인디언의 유전자 형성에 대한 유전적 기여를 밝혀 냈습니다 ( Lazaridis et
al., 2014 ; Raghavan et
al., 2014 ).
유라시아 유전 환경에서의 실질적인
변화는 청동기 시대(약 3천 ~ 2천년)에 일어 났는데, 이는 유럽으로 대규모 인구 이동과 인규 교체를 포함하는 주요 변화였습니다
( Allentoft 외
2015 ).
후기 Proto-Indo-European와 관련된
Yamnaya 문화는 남서부의 시베리아 우랄 지역과 남동부 유럽의 Pontic 대초원 지역에서 발생했습니다 ( Allentoft 외
2015 ; Haak
외 2015 ; Jones 외
2015). 동유럽과 CHG(=코카서스 수렵 채집인) 그룹으로 그들의
기원을 추적한 Yamnaya Pontic Steppe인들은 유럽의 신석기 농경인과 그 문화를 대거 교체합니다. 결과적으로 유럽의 많은 지역에 걸쳐 청동기 시대인들은
신석기 농경인과 비교하여 훨씬 많아 지게 됩니다 ( Haak 등
2015 ). 동시에, 초기 동-서 유럽 사람들은 청동기 시대에
접촉했습니다. 독일의 후기 신석기
농부들은 약 75%가 Yamnaya 사람들로 교체되었다 (Haak et al.,
2015). 이러한 이주 현상은 유럽과 시베리아의 현재 인구 구조에
영향을 받았지만 그 세밀한 내부 세부 사항은 크게 불분명하였다. 시베리아 원주민 집단의 유전자 구성과 유럽 및 동아시아
인구와의 유전적 연관성에 대해서는 거의 알려지지 않았다. 게다가 이전의 연구들은 고대 시베리아 토착민 그룹의
게놈들, 특히 Mansi, Khanti 그리고 Nets 사람들의 서부 시베리아 인구에 대한 고대 유라시아 인들의 유전적 기여도를 알아
내지 못했다.
시베리아는 일반적으로
서부및 동부 지역으로 세분화됩니다. 서쪽 시베리아의 영토는 서쪽의 Trans-Uralic
지역에서 동쪽의 예니세이 강까지 이어집니다. 이 지역의 다른 여러 원주민 뿐만 아니라 Mansi,
한티(=Khanti)및 네네츠(Nenets) 의 서부 시베리아 인종 그룹은 크게 Uralic 언어를 사용하며 Ugric, Finno-Permic
및 Samoyedic 언어 그룹으로 세분화됩니다 ( Supplemental
Fig. S1). 서-시베리아인 한티와
만시는1,300만명인 중유럽의 헝가리인들과 우그릭 언어 그룹의 하나입니다.
북서부 시베리아 네네츠 사람들의
언어는 우랄어 (Uralic), 사모예어 (Samoyedic) 계통에 속하며, 이로 미루어 보면,네네츠 사람들과 다른 서부 시베리아 인구들 사이에도 유전적
공유가 되어 있음을 시사합니다. 이런 지리적인 차이에도 불구하고 몇몇 유럽 사람들은 서부
시베리아 인이 말하는 Ugric 언어와 관련된 언어를 사용합니다. 이들은 전통적으로 북부 및 북동부 유럽 지역을 차지하는
Komi, Karelians, Veps, Saami 및 Finns가 포함됩니다. 그들의 언어는 Permic 및 Finno-Volgaic
언어 그룹에 속하며 다른 Ugric 언어와 관련이 있습니다 ( Supplemental
Fig.
)
동부 시베리아 사람들은
시베리아 타이가 숲 지역에서 서쪽의 예니세이 강과 동쪽의 오호츠크 해 사이에 정착했다. Evens, Evenks 및 Yakuts가 차지한 지역은
시베리아 동부 지역의 대부분을 차지합니다. 알타이족의 전통적인 영토는 시베리아 타이가, 카자흐스탄의
대초원, 몽골의 고비 사막의 교차점에서 아시아의 중심에 있습니다. 동유럽의 볼가 (Volga) 지역에 정착 한
Kalmyks족은 중국 북서부 지역의 쯔가리아 (Dzungaria)에서 17 세기에 이 지역으로
이주했다 ( Erdeniev
1985 ). 칼 시크 (Kalmyk)및 알타이 (Altayan) 언어와
함께 동 시베리아 원주민의 언어는 크게 알타이 (Altaic)로 분류되며 몽골, 퉁구스, 투르크 족으로 세분화됩니다.
( 보충 그림
1 ).
이 연구의 초점인 인구 표본 간의 상대적 이격 거리를 추정하기 위해, 처리된 게놈을 '주 구성
요소(principal
components)'에 사상(寫像) 했다. 해당 게놈이 이
그룹의 집단을 대표하는 훌륭한 샘플로 선택한 후, 게놈은 클러스터링 되어 시퀀싱
처리되었다
그림1. <유럽및 시베리아 인구의 지리적및 유전적
친연성. (A) 시베리아, 서부 러시아 및 동 유럽지도는 이 작업의
일환으로 개별적으로 염기
서열 분석된 결과인 위치를
보여줍니다.지도의 색상 영역은 전통적으로 해당 인구가 차지하는 지리적
영역을 보여줍니다 ( Peoples of
Russia 1994). 유럽과 시베리아 사이의 자연 경계인 우랄 산맥은 파란
봉우리로 표시됩니다. 서쪽 시베리아의
소수 민족인 Mansi, Khanty 및 Nenets 사람들은 서쪽 시베리아의 우랄 산맥에서 시베리아 동부의 예니세이 강까지 뻗어있는 타이가의
광대한 지역에 분포합니다. Komi,
Karelians 및 Veps를 포함한 북동부 유럽 인구는 우랄 산맥의 서쪽 지역을 차지합니다. Altayans, Yakuts, Buryats, Evens
및 Evenks를 포함한 동부 시베리아 인구는 서쪽의 예니세이 강과 동쪽의 오호츠크 해 사이의 시베리아 타이가 지역을 차지합니다.
Kalmyks족은 역사적으로 중국 북서부의 지역에서 낮은 볼가 지역에
이동하였읍니다.. (B) 시베리아와 유럽에서 온 27 명의 인구를 대상으로 892
명의 현대인을 대상으로 한 주성분 분석 (PCA). PCA는 137,637 개의 공통 SNP 세트를 사용하여
수행되었습니다. 시퀀스된 샘플은
별표를 사용하여 표시됩니다. DNA
microarray를 사용하여 genotyped 샘플은 점으로 표시됩니다. >
PCA 플롯은 첫 번째 주성분에
따라유전적 변이의 6.4% 변화로 집단의 주요 분화를 포착합니다. 첫 번째 주요 구성 요소는 유라시아의 서쪽에서 동쪽으로
기울기에 해당합니다. 두 번째
주성분은 북쪽에서 남쪽으로, 특히 시베리아인들 사이에서 인구의 확산을 포착하여 유전적 변이를 1% 정밀도로 포착했다. PCA 플롯은 시베리아 집단 간의 상당한 유전적 차별을
나타낸다. 이것은 시베리아 인구가
다른 개체군 (예 : 매우 인구 수준의 차이가 매우 적은 수준의 유럽 인구 집단)에 의해 대표되지 않는 중요한 유전적 다양성을 갖고 있다는 것을
의미한다.
동서 시베리아인들 사이의 혼혈
양상
다음으로 우리는 TreeMix ( Pickrell and
Pritchard 2012 )를 사용하여 전체 유전체 염기 서열 분석 데이터에서
유추된 3천 백만개 게놈 전체 상 염색체 SNP 세트를 사용하여 인구 유전 관계 및 인구 집단 혼혈 트리 기반 모델을
구축했습니다. 이 모형은 2 개의
초기 분기점을 따라 유럽과 아시아 인구를 배치했다 ( 그림
2 ). 또한, 서부 시베리아 만시 (Mansi), 한티
(Hanty), 네네츠 (Nenets) 그룹은 다른 동유럽 인구와의 친연성을 지닌 초기 분화를 형성했다. Even, Evenk, Buryat 및 Yakut 그룹을
포함한 대부분의 동부 시베리아인들은 동아시아 인구와 더 관련이 있는 별도의 계보를 형성했습니다.
그림2. <현대의
시베리아, 아시아, 아메리카 원주민, 유럽 및 아프리카 인구의 유전적 관계. Autosomal TreeMix 혼혈 그래프는 아프리카,
유럽, 시베리아 및 미국에서 온 44 명의 개체를 대상으로 한 전체 게놈 시퀀싱 데이터를 기반으로 한다. X축은 유전적 drift를 표시하는데, 유효 인구 집단
크기인 Ne에 비례한다. 혼혈(admixture) 이벤트는 화살표로 표시되며 혼합 강도는
백분율로 표시됩니다. 나머지는 보충 그림 S5에
나와 있습니다.>
TreeMix 모델은 또한 몇 가지 중요한 혼합 이벤트를
나타냅니다. 특히 서부 시베리아
조상의 43%(95% CI: 38%-47%)는 현대의 에벤키 사람들과 가장 관련이 있는 고대 동-시베리아인과의 혼혈에 기인한 것으로 나타났습니다. 북서부의 시베리아 네네츠 사람들은 동부 시베리아인인 현대의
에벤키인과 밀접한 관계가 있는 상태에서 추가적인 혼혈의 증거를 보여 준다. 우리는 네네츠의 조상 중 38%(95% CI:
31%-46%)가 이 혼혈에서 기인한 것으로 추정했다. 이러한 예측을 더 확인하기 위해,
우리는 서 시베리아와 동 시베리아
그룹 간의 공유된 대립 유전자를 테스트하기 위해 D-statistics ( Durand et
al. 2011 ; Patterson et
al.보충적
그림. S7A-C, S8A-C,
S18 )을 테스트하였다. TreeMix 추론과 더불어, 우리는
D-statistics 테스트 ( Supplemental
Figures S7A-C, 8A-C )에 근거하여 Mansi와 Evenk, Khanty and
Evenk, Nenets, 그리고 Even 사람들 간에 유의미한 유전적 친연성을 관찰했다. 이러한 관찰은 근대 서부 및 동부 시베리아 인구의 조상 간
고대 혼혈이 있음을 입증한다.
특히,
TreeMix는 Mansi, Khanty 및 Nenets와 아메리카 원주민인 Andean High-landers의 조상 간에 공통된 유전적
혼혈이 있음을 추론했다. 이러한
혼혈 결과는 서부 시베리아 조상의 6%(95% CI: 4%-8%)를 차지합니다. 아래에서 논의되는 바와 같이,이 혼혈은 서부 시베리아 인과
인디언의 '고대 북 유라시아인(=ANE)'들에 대한
유전적 연결로 설명될 수 있다.
계통 발생도는 조상
계통의 분기 시간에 대한 정확한 정보가 부족한 단순화된 인구 통계 모델을 제공합니다. TreeMix 브랜치의 길이는 드리프트 매개 변수의 단위에
비례하며 ( Pickrell 및
Pritchard 2012 ) 인구 분산의 시간과 직접 관련 될 수
없습니다. 또한 분기 길이는 인구
병목 현상의 영향을 받을 수 있으므로, 인구 분산 시간의 추론을 복잡하게 만들고, 모델의 해석 가능성을 제한합니다. 모집단 간의 발산 시간을 보다 직접적으로 추정하기 위해
여러 순차적으로 Markovian coalescent (MSMC) 분석을 사용했다 ( 그림 3A ; Schiffels
and Durbin 2014). 우리는 두 개체군 사이의 관련 교차 유착률이 이 분리
시간의 대용으로 0.5가 되는 시간을 지정했다 ( 그림 3B). MSMC 방법은 각 개체군이 단 한 명으로만 표현될 때
10,000 년 이상 된 사건에 특히 주목합니다 ( Schiffels
and Durbin 2014 ). 10,000년 전보다 더 최근의 추정치는 더 많은 개인이
분석에 포함되지 않는다면 덜 정확합니다.
그림 3.
그림 3.
<인구 분기 시간에
대한 MSMC 분석. (A) 5 명의 참고 개인 (프랑스, Andreapol
러시아어, Mansi, Evenk 및 Han) 각각에 대해 분기 시간을 계산했습니다. 기준 인구
모집단은 상단 (오렌지색 둥근 사각형 안에 있음)에
표시됩니다. 각 기준 열은 기준
인구와 다른 참조 인구 사이의 MSMC 분리 시간을 보여줍니다. 다른 참조 인구는 가장 최근
(=상단)에서, 가장 오래된 (하단)으로 분리 시간에 따라 분류되었습니다 . 분리 시간은 왼쪽에 표시됩니다. 각 그래프의 측면. 유사한 시간을 가진 인구가 함께 그룹화
되었습니다. Andreapol; (Ust) Ustyuzhna; (Ob) Objachevo. 10,000 년 미만의 MSMC 분리 시간은 noisy
하므로 이러한 분리 시간과 해당 모집단은 회색 글꼴로 별도로 표시됩니다. ( B ) Mansi와 다섯 개의 다른 개체군 (Nenets,
Evenks, French, Andean Highlanders, Dinka) 사이의 차이에 해당하는 MSMC cross-coalescence
그림. 중간 분리 값
( y = 0.5, 주황색 선)은 모집단 분화 시간의 대용으로
사용되었다.>
우리의 MSMC 분석은 서양 시베리아의 선조 집단이 현대의
에벤키 인구와 가장 관련이 있는 고대 집단과의 혼합을 겪은 후에 Mansi, Khanty 및 Nenets가 비교적 최근에 분리되었음을 제시한다
( 그림
2 ). 이 혼혈은 Mansi와 Evenk 인구 사이의 예상 분리
시간을 기준으로 ~6.8kya로 계산 될 수 있습니다. 이 시간은 대략적인 것으로 주의해서 해석해야
합니다. 그러나, 인구의 상대적인
순서 ( 그림
3A)는 여전히
상대적인 유전적 유사성을 해석하는데 유익합니다. 이 추정치를 더 확인하기 위해 인구 당 두 명의 개인을
사용하여 보다 계산 집약적이지만 보다 정확한 8-haplotype MSMC 분석을 수행했습니다. 이 분석의 결과는 서부 시베리아 인구에 동 시베리아 섞인
시간 9.9kya의 비슷한 추정치를 제공했다 ( 보충 그림
S9 ). 따라서 동부와 서부 시베리아 인구는 지난
5,000~10,000 년 사이에 서로 접촉한 것으로 보인다.
현대 유럽인과 시베리아 인의
관계
계통수 발생
모델은 거의 모든 현대의 유럽인들 (중국에서 약 400년 전에 유럽으로 이주한 Kalmyks를 제외하고)을 단일 지점을 따라 배치했으며, 유럽
내에서 인구의 동서 분포를 모방한 분화 패턴을 가졌다 ( 그림 2
).서양과 동유럽
인구가 유전적으로 비슷하다는 PCA
분석 결과 ( 그림1B)를 보면, MSMC 결과에 따르면 프랑스와 동유럽
인구의 분리 시기는 2.2kya (프랑스 - 카렐리아) ~ 7kya ((프랑스, Komi
Objachevo). 대조적으로,
프랑스인과 시베리아인 사이의 최소 분리 시기는 12.5 kya (프랑스-한티)였으며, 이는 유럽인들 사이의 분리 시간을 크게
웃돌았다.
우리의 상 염색체 결과에 대한
추가 자료를 제공하기 위해 Y 염색체의 일부분 안의 SNP와 개체군 특이 하플로 그룹의 추정 된 분기된 시간을 분석했다 ( 그림 4A). 우리의 Y 염색체 분석 (방법, 보충
정보 "Y 염색체
및 mtDNA 분석"절 )은
시베리아 인구에서 동유럽및 북유럽으로의 유전자 이동의 증거를 보여주었습니다. N-haplogroup 인 Y-Chromosome
clade는 고대 시베리아 인구와 관련이 있다 ( Fu et al.,
2014 ). N 하플로는 현대의 시베리아와 동유럽에서 일반적이다
( Malyarchuk
et al., 2004 ; Lappalainen
et al., 2008 ; Mirabal et
al., 2009).) 그러나 서유럽인은 이것이 없다. 우리는 시베리아 (Evens, Evenks, Mansi,
Khanty)와 북동부 유럽 인구 (Veps와 Komi) 사이에서 ~5.3-7.1 kya에서 N1c1 Y 염색체 haplogroup의 확장을
관찰했다. 이것은 N1c1
haplogroup가 상대적으로 최근에야 유럽에 도달했으며 서유럽 인구로의 확산은 제한되었음을 시사한다.
그림 4. <Y 염색체 및 mtDNA 계통 발생과 haplogroup divergence
times. ( A ) Y-Chromosome tree
topology는 Poznik et
al.이 정의한 신뢰도가 높은
지역에서 12,394 개의 다형성 부위를 사용하는 MEGA 도구 ( Tamura et al.
2011 ) 에서 구현된 최적 가능성 접근법을 사용하여
계산 되었습니다 (2013) . X 축으로써 행한 다음 하플로 분화 시간을 나타내며, 수천년
단위로 표시된다. (1
KYA=1,000년이다). 이 나무는 30 명의 개체군의 54 명의 현대인의 자료를
사용하여 만들어졌습니다. 분할 시간은
각 가지 (방법)에 할당되도록 유도된 대립 유전자의 수에 따라 계산되었고, 분할은 하플로 분화 시간을 반영하기
위해 x- 축을 따라 배치되었다. 터미널 분기는 X = 0에서 끝납니다. 7 명의 고대 개체가 각 지점에 할당된 높은 신뢰도의
SNP를 가진 SNP 공유 패턴에 따라 하플로 계통수에 배치되었습니다. 고대 인골(오렌지 라인)의 지점은 각 고대인이 살았을 때
종료됩니다. Haplogroups는 ISOGG Y 염색체 SNP 지수를
기반으로 할당되었고 나무의 오른쪽에 표시되었습니다 ( International Genetal Genealogy
2014 ). 이 작업의 일부로 시퀀스 된 개인은 빨간색으로
표시됩니다. (B) mtDNA 계통 발생은 Y-Chromosome tree와
유사한 방식으로 MEGA 도구를 사용하여 구축되었다. 이 나무에는 70 명의 개인과 34 명의 인구와 8개의 고대
게놈 데이터가 포함되어 있습니다. (C) Y 염색체 계통 발생 (왼쪽)과 mtDNA 계통 발생 (오른쪽)에서 haplogroups의 분기 시간. 발산 클레드가 왼쪽에 표시됩니다 ("Split"열). 신뢰
구간은 포아송 (Poisson) 모델을 사용하여 계산되었습니다.>
흥미롭게도,
mtDNA 하플로
분화( 그림 4B)로
구축된 계통수는
5-10kya로 시베리아인과 북동부 유럽인들 사이에서 뚜렷하게 구별되는 군집을 보여주지 못했다.그러나 우리는 mtDNA 계통수가 매우 많은 다양성을
가지고 있다는 것을 알고 있다. 그리하여, 우리는 서부 시베리아와 북동부 유럽인 사이의
마지막 7.000년 동안 발생하는 유전자 흐름을 알아 내기 위해 D-통계 시험을 추가해 분석하였다 ( 그림 5A, B, 보충 그림 S7G,
K, L, S8G, K, L, S13A , B ). N1c1 haplogroup 확장과 일치하여, Mezen
러시아인, Veps, Karelians 및 Komi를 포함한 북동부 유럽인은 Nenets, Evens, Yakut 및 Khanty와 같은
시베리아 집단과 통계적으로 유의한 혼혈 신호를 보였다. 이러한 혼혈은 Ustyuzhna와 Andreapol
Russia와 같은 동부 유럽의 남반구 인구와 벨로루시인 ( Supplementural fig. S7H-J,
S8H-J ) 중 훨씬 더 약하며 시베리아 혼혈은 특히 북동부 유럽인들 사이에서 강하게
나타났다.
그림 5. <D-statistics를 이용한 혼혈의
분석. 유전적 유사성은 이전에 정의된
대로 D- 통계 테스트 D (P1, P2, P3, O)를 사용하여 결정되었다 ( Durand et al.
2011 ). (A) Mezen 러시아인 친연성; (B) Veps 동질성; (C) Ust'-Ishim 동질성; (D) Mal'ta 소년 (MA-1)과 Afontova Gora
(AG-2) 친연성; (E) Ire8 및 Ajv52 친연성. 다른 모집단에 비해 모집단 X (오렌지색)의 유사성은
ADMIXTOOLS ( Patterson 외.
2012 )를 사용하여 계산되었으며, D열 아래에 표시되었습니다. 혼혈의 통계적 유의성은 z-점수 및 Z열 아래에
표시됩니다. 녹색 글꼴은
P1과 P3 ( Z >
3) 사이에 중대한 혼혈을
강조하기 위해 사용되었습니다. 파란색 글꼴은
P2와 P3 사이의 유의한 혼혈을 강조하기 위해 사용되었습니다 ( Z <-3). 아프리카 산은 모든 테스트에서 아웃 그룹으로
사용되었습니다. Andreapol; (Ust) Ustyuzhna; (Ob) Objachevo. 다른 아웃 그룹 (Chimp, Mandenka,
Mbuti)을 사용한 D- 통계 결과는 보충 그림
S13 및 S17에 나와
있습니다.>
동부 시베리아인들과 동아시아인들 간의 유전적
유사성
TreeMix 결과를 토대로 우리는 동부 시베리아 사람들이 유럽인보다 아메리카 원주민과 동아시아 인구에 강한 유전적
친연성을 가지고 있음을 관찰했다 ( 그림
2 ). Buryats, Yakuts, Evens, Evenks와
같은 동부 시베리아 인구는 동아시아인 Han, Dai 및 Sherpa ( 그림
2 , 보충 그림 S3,
S4 )에서 분기된 후에 다른 하위 계보를
형성합니다. MSMC를 사용하여 우리는 동부 시베리아인과 동아시아인
사이의 분리가 8.8-11.2 kya (Evenk-Han 및 Evenk-Sherpa 분리 시기 추정치를 기준으로 함) 사이임을 확인했습니다
( 그림
3A ). Eastern Siberian 상 염색체 클레드의 존재와
동아시아 인구에 대한 친연성은 다중 TreeMix 모델에 의해 잘 지지
된다 (도
2 2,6,6 ,7;7 ; 보충 그림
3 ), 100 % 부트
스트랩 복제 (그림
6 ).
따라서 우리의 연구 결과는 동 시베리아 계보와 다른
동아시아 그룹의 분리를 지지하며, 우리는 이 분리 현상을 10kya로 추정했다.
그림6.
<Ust'-Ishim과 현대 인간의 유전적 관계. 7개의 혼혈이 발생한 TreeMix 모델에는 고대 시베리아
Ust'-Ishim이 포함되어 있습니다. 파란색 원은 분기 event가 일어난 것으로 100 부트 스트랩을
사용하여 조사된다. 분기점을 지원하는 트리의 백분율이 표시됩니다. 나머지는 보충 그림 S14에
나와 있습니다.>
그림7.<고대 북 유라시아 인 MA-1과 AG-2와의 유전적 관계. Autosomal TreeMix는 고대의 7,000년 된
La Brana (스페인)와 고대 시베리아인 MA-1및 AG-2의 낮은 커버리지 게놈을 포함하는 그래프입니다. 이 그래프의 나머지 그림은 보충 그림 S16에
나와 있습니다. TreeMix 모델은 MA-1과 AG-2로 대표되는 ANE
계보에서 41%의 혼혈 비율 (95% CI: 36%-45%)을 Andean High-landers가 대표하는 아메리카 인디언으로, 43%를 혼혈
(95%CI : Evenen과 현대인 네네츠와 관련된 인구 사이의 26% (95% CI:16%-37%)의 서부 시베리아인 (Mansi and
Nenets)의 공통 조상과 관련한 인구와 38 % -47 % . 또한, 서부 시베리아인들과 ANE는 함께 분류되어 그들의
유전적 역사를 공유합니다.>
Kalmyks족은 중국 북서부에서 유래 되었으나 나중에는
남동부 유럽으로 이주했다. 따라서
다른 개체군, 특히 다른 시베리아 집단과의 유전적 친화력을 결정하는 것은 옳을 것이다. 시베리아와 동아시아에서 Kalmyks와 그들의 역사적
지리적 이웃들과의 관계를 명확히 하기 위해 우리는 추가적인 D 통계 분석을 수행했다. Kalmyks와 알타이 족은 모두 동부 아시아인들 (D 통계치 =
0.13-0.146 , 보충 그림들 S7D,
E, S8D, E )을 포함하여 동부 시베리아인들과 비교적
가깝다 (D 통계 =
0.11-0.125 ). 그러나
Kalmyks와는 달리 Altayans는 현대 유럽인과 서양 시베리아인 모두와 관련이 있다고 예측한 또 다른 알려지지 않은 인구에 대해 그들의
조상의 ~ 37 % (95 % CI: 31%-43%)를 추적했다 2).
고대 게놈에 대한 링크
현대 시베리아 인과 Ust'-Ishim의 유전적
유사성
(고대의 인구
통계학적 이벤트를 발견하고 오늘날 개인과 고대의 조상 집단 간의 유전 링크를 표시하려면, 우리는 현대의 인간에서 60개 게놈 비교 표
1 , 보충 표
S2를 46개 고대의
게놈 보충 표
S3 45,000- 포함), 2만 4천년 된 시베리아 '말타
소년(MA-1)'과 17,000년 된 시베리아 '아폰토바 고라 (AG-2)'으로 구성된 시베리아의 이스트 인 ( Fu et
al. 크라스 노야르스크
지역 ( Raghavan 외
2014 ).
서쪽
시베리아에서 Ust'-Ishim 개체의 뼈가 발견된 것을 감안할 때 Ust'-Ishim은 현대 시베리아 인구와 관련이 있을 것이라고
추론했습니다. 실제로,이 가설은 잘
TreeMix 모델 의해 지원되었다.(그림
6 ). 우스트 이쉼 조상은 시베리아와 동아시아의 공통 조상
공유 38% (28-48%, 95% CI) 추적 관찰 (그림 6, 보충
데이터S14)
된다.
이와
관련해서, D-통계 분석 ( 그림5. C; 보충 데이터 S13C,
S15)는 Ust'-Ishim이 동양인이나 시베리아인들, 특히 동부 시베리아인들과 현대 유럽인들보다
유전적 친화력이 더 높음을 보여 주었다.이것은 또한 Y-haplotype 분석에 의해
뒷받침되었다. 이전에는
시베리아 Ust'-Ishim( 푸 et al.
2014 )
일반적인 'R' (유럽에서 매우 흔한 Y-염색체 계통군), 하플로 그룹의 'Q'를 하플로 그룹 조 인 'K2' Y-염색체 계통군에 속하는 것으로
생각되었다. (시베리아와 북동부 유럽에서는 공통). O 집단 (동아시아에서 공통적인 집단)으로 분류된다. 그러나 Ust'-Ishim의 NO 분기 (약 7,690,182 Y 좌표 hg19)
(특정 Y 염색체에서 단일 SNP 유래한 것으로 관찰. 보충 데이터.
S12). 이 branch는 동양인 뿐만 아니라 시베리아인들과 동북
유럽 그룹들과 관련이 있습니다. 따라서 이전 주장과는 달리, Ust'-Ishim의 haplogroup은 더 분기된 K2 clade보다는 가장 최근의
NO clade에 속할 가능성이 큽니다. 이러한 증거를 바탕으로 우리는 Ust'-Ishim이 현대 유럽인이나 아메리카 원주민보다
동아시아와 시베리아 인구에 더 많은 조상을 기원했다고 결론을 내립니다.
서 시베리아 인은 ANE(=Ancient North
Eurasian) 및 동부 시베리아 인의 후손입니다.
우리의 계통수
모델은 ANE(=고대
북유라시아인) MA-1과 AG-2를 서부 시베리아인들 (Mansi and
Nenets)과 그룹화하여 ANE와 근대 서부 시베리아인 사이의 유전적 연관성을 제안했다 ( 그림
7 ). 또한 TreeMix는 아메리카 인디언인 Andean
High-landers의 조상 중 41% (95% CI: 36%-45%)가 ANE 혈통에 기인한 것으로 추정했다
( 그림
7 ; 보충 그림
S16 ). 이것은 Paleo-Americans가 상 염색체 게놈의
~42%를 MA-1과 관련된 고대 유라시아 계통 ( Raghavan et
al. 2014 )으로 추정한 이전 보고서와 일치 합니다. 우리는 현대인이 동아시아 한 개체보다 MA-1 또는
AG-2와 가깝게 된 대립 유전자를 공유하는지 평가함으로써 D-통계를 사용하여 이러한 예측을 시험했다 ( 그림
5디; Supplemental
Fig.17A ). Mansi, Khanty, Nenets 및 Native
Americans 모두 MA-1및 AG-2와의 유의미한 유전적 친화성으로 입증된 매우 강한 ANE 조상을 가졌습니다. 따라서 서부 시베리아인들은 아메리카 원주민과 공통된 ANE
조상을 공유합니다. 서쪽 시베리아
사람들의 남은 조상은 동 시베리아 사람들과 관련되기 때문일 수 있습니다. TreeMix 모델 ( 그림
7 )은 Mansi 조상의 43%가 동 시베리아의
Evenks and Evens와 관련된 인구와의 혼혈에서 파생되었고, 57 %는 ANE 조상에 기인한 것으로
추정했습니다. 그러므로 Mansi
종의 게놈은 시베리아 ANE 사람들과 동 시베리아 인구에서 상당한 비율로 거의 같은 조상을 보유하고 있습니다.
고대 유럽의 시베리아
관련 조상을 조사하기 위해 현대
러시아의 사마라와 카렐리아 지역의
홀로세 동유럽 사냥꾼 ( 하크 외
2015 )과
스웨덴의 모탈라 지역 (6.6-8Kya)의 SNP 데이터를 분석했다. 더 최근의 Bronze
Age Yamnaya 샘플 (4.7-5.3 kya). D-Statistics는 Mansi와 동유럽의 거의 모든
수렵 채집인들 간에 강력한 혼혈을 보였다 ( Supplemental
Fig. S21), 특히 Samara와 Karelia 샘플의
경우. 시베리아 동부와 서부 유럽
수렵-채집인들 간에는 약간 약하지만 중요한 유사성이 있었습니다. 앞서 언급했듯이, 동 시베리아 인구와 서부 시베리아 인구가
분리된 시기는 약 6.8-9.9 kya로 추정됩니다. 동 시베리아인들과 동유럽 수렵 채집인들의 약한 친연성
정도는 Mansi와 관련된 혼혈을 통해 고대 유럽 인구로 스며든 Mansi의 동 시베리아 조상에 기인할 수 있습니다. Yamnaya 표본은 또한 Mansi와 Evens와
통계적으로 유의한 혼합을 보였으나 Yamanaya 문화 표본의 Eastern hunter-gatherer 조상 구성 요소의 희석을 나타내는
Samara HG와 비교하여 약간 약했다.
시베리아 인들은 동유럽 수렵 채집인 외에도 스웨덴의
Ire8과 Ajv52 (2005 년 Skoglund
외 )의
5,000년 된 수렵 채집인과 PWC (Pitted Ware Culture)의 조상의 일부를 공유했다. Ajv52와 특히 Ire8은 서양 쪽 시베리아 인
Mansi와 강한 혼혈를 보였으며 ( 그림
5 E, Supplemental
Fig. S17B ) 밀접한
관련이 있는 Yamnaya 문화와 마찬가지로 Mansi 관련 인구와의 혼합으로 ANE 혈통이 강할 가능성이 있음을
시사한다.
동부 시베리아 인과 Saqqaq 문화의 고대 인물 사이의
조상 공유
우리의
데이터는 또한 Greenland의 4,000 년 된 Saqqaq 개체 ( Rasmussen et
al. 2010 )의 게놈 내에서 동 시베리아 조상의 증거를 보여
주었다. Saqqaq 게놈을 가진 Tree 모델은 Saqqaq이 동
아시아 인과 동 시베리아 인과 관련이 있고, 그 조상의 12 %가 현대 에반스와 관련된 인구에 기인한 것으로 나타났습니다
( 보충 그림 S19,
S20 ). 검출된 유전자 흐름은 D-statistics
( Supplemental
Fig. S7F, S8F )에 의해 강력하게 뒷받침되었다. 또한, 미국 안데스의 조상 중 18 %가 Evens
( Supplemental
Fig. S19). 이것은 Evens and Evenks의 조상으로부터
Altayans에 이르는 상당한 유전자 흐름과 함께 Evens에서 Saqqaq로의 신호가 시베리아 인 (동부 시베리아 인)의 조상과 초기
아메리카 대륙 이주민 사이에서 일어난 고대 혼합물의 결과일 수 있음을
시사합니다.
토착 시베리아 원주민의 게놈은 유라시아와 유럽의 인구
역사와 인구를 형성하는 많은 중요한 고대 사건들을
촉발시키는 유전자 요람(=harbor)이라는 중요한 증거를 가지고
있다. 시베리아는 30개 부족
종족 집단의 본거지이지만, 지금껏 이런 사실들은 간과되고 연구되지 않았습니다. 이 연구에서, 우리는 여러 시베리아 및 동유럽 인구의
유전적 역사를 조사하고 고대와 현대 인구에 대한 유전적 연결을 분석했습니다. 우리는 인구와 유전 변이가 북 유라시아 전역에
퍼지는 지리적 인구 확산 모델
( 그림
8 )의 형태로 본 연구 결과를 다음과 같이
요약했다.
( 필자 멘트: 참고로 이 모델은 그냥 개념적인 것으로 봐야 한다. 4.7만년 전의 우스트이쉼 유골은 순다
대륙에서 역확산하여 우랄 평원에 진입한 것이다. 이 역확산 루트로 필자는 시계 반대 방향으로 '툰드라 하이웨이'를 주장한다. 여기서는 시계
방향으로 인도를 경유하여 유럽에 진입하는 것으로 해석하였지만, 조만간 필자의 주장대로 바뀔 것이다. 왜냐하면 지난 2년간 발표된 고이골 유전자 분석 결과는 하나 같이, 필자가 주장하던
듣보잡 가설들을 하나씩 입증해 주고 있기 때문이다. 물론 처음에는 이런 결과가 나왔을 때도 차마 이를 믿지 못하고 다르게 해석하다가, 결국에는
필자가 언어학 기반 제시한 모델로 귀결되었기 때문이다. 이제 남은 마지막 퍼즐을 맞출 고리가 바로 '툰드라 가설'이다. 이와 같은 고인골은
3.7만년전의 모스트바 남쪽의 코스텐키 유적의 유골과 벨기에의 고예트 유골에서도 일관되게 보여 주는 현상이다. 이들의 기원은 '동남아'에서
기원한다. 이들은 3만년전까지 전 유럽에 퍼져 유럽의
고대수렵채집인을 형성하는데, 현대 유럽인들은 이들과 사촌 쯤 되는
데서부터 현재 유럽인들이 시작되었다고 보는 것이 현재까지 분석 결과이다. 이들은 3만년전부터 Gravettan 구석기인들로 바뀐다. 이
Gravettan인들은 아마도 새로 남방에서 유입된 사람들로 보인다. 그러나 1만9천년부터 시작된 최대 빙하기때 이들은 사라지고, 최대 빙하기를
거치면서, 다시 3만년전의 Aurignac 구석기인들이 피한지에서 혼혈을 거친 다음 재
등장한다. 이들이 'WHG(=서유럽수렵채집인)'을 형성한다. 기원전 6천년 쯤부터 유럽에서
농경이 시작된다. 이때는 홀로세 최적기(Holocene
Climate Optimum, HCO)라 하여, 시베리아가 살기 좋은 때였다. 그러나 이 '신석기 농경인(=Neolithic Farmer)'들도
따지고 보면, 주류는 Aurignac인들이 자체 분기하여 형성된 WHG 사람들이 유전적 분기한 것이다.
이들은 유럽으로 재 진입을 하고, 현지의 수렵채집인(=WHG)들과도 나중에는 동화를 한다. 기원전
3천년부터는 어떤 이유인지는 몰라도, 신석기 농경인들은 흑해 북단의
볼가강 지역의 얌나야인들로 유럽의 남성은 독일의 경우에 75%까지 교체가 된다. 그러나 이들은 2,4만년전 바이칼의 말타 부렛 고인골의 후예이고, 이런 증좌는 1.7만년전의 알타이
지역의'아폰코바 고라' 고인골 유전자와 유전적 연속성을
지닌다. 다시 말하면,
3~4만년전에 북방 툰드라에서 유럽인들이 유입되고, 이들이 자생적으로 자라다가, 다시 기원전 3천년 경부터, 대규모 인구 이동이 시베리아로부터
있어, 현재 유럽인구 구성이 완료된다. 따라서 이런 관점에서 본다면, 아래 그림은 뭔가 이상하다. 아마도 백계 러시안의 종주권을 주장하고
싶어하는 의도가 깔려 있는 것 같다. )
그림8.<지리적 인구 확산 모델. 염색체 Y, MSMC, TreeMix 및 D- 통계 분석을
기반으로, 대략적인 분기 시간을 보여주며, 유럽, 동아시아 및 시베리아의 1차 유전적 계보(=클레드)의 형성을 반영하는 유전자 흐름의 근사
모델. 백분율은 다른 계보에서 각
혈통 형성에 기여한 조상의 비율을 나타냅니다. Mansi, Khanty 및 Nenets의 공통 조상은
말타(MAE-1),아폰토바고라(AG-2) 및 아메리카 원주민과 관련된 ANE(=고대 북 유라시안) 에서 57%가 기원하고, Evens
and Evenks와 관련된 인구와의 혼합에서 43%가 기원합니다. 아메리카 인디언들은 ANE(=고대 북 유라시안)에 그들의 조상의 42%를 물려 받고, 동 시베리아인과 동
아시아인의 공통 조상에게 58%를 물려 받습니다. 그림에서 45kya(=4.5만년)는 Y 염색체 'Q'와
'R'이 'NO'와 분리됨을 나타냅니다. 33 kya는 Y- 염색체 'Q'와 'R' 계보가 서로
분리되고, 또한 mtDNA 'N2'와 'W' 계보의 분리를 나타냅니다. 17 kya는 Mansi와 Andean이 분리됨을
나타냅니다. 44 kya는 NO
계보로부터 Ust'-Ishim(=Kx(LT))의 haplogroup의 분리를 나타낸다. 19 kya는 Han 과 Andean의 분리
됩니다. 10 kya는 Han에서
East Siberian의 분리를 나타냅니다. 5.3-9.9 kya는 Evenk-Mansi, MSMC
분리(6.8-9.9 kya)와 N1c1 Y-haplogroup (5.3-7.1 kya)의 확장을 나타냅니다. 이 그림에서, 유럽인은 이 연구에서 Komi, Veps, Karelians, Russia,
Belarussians 인구 집단의
총칭입니다 :.>
지금까지 분석된 현대 및 고대
유라시아 게놈의 대부분은 북유럽과 서유럽, 유라시아 대초원 지역 및 남부 시베리아에서 수집된 것이다 (Skoglund et al.,
2012; Fu et al.,
2014 ; Gamba et
al., 2014 ;
Lazaridis et al 2014; Raghavan et
al. 2014 ; Allentoft et
al., 2015 ; Haak et
al. , 2015 ). 이와 대조적으로, 여기서는 시베리아 및 러시아의 유럽 지역
타이가 (Taiga) 등에서 샘플을 수집했습니다. 우리는 이
지리적으로 떨어진 별개의 그룹에서 현대와 고대 개인 사이의 유전적 친연성을 뚜렷하게 관찰하여, 동북 유럽 및 시베리아 인구의 유전 구성을
형성하는데 있어, 시베리아를 가로 질러 아시아 및 유럽의 대규모 인구 이동을 있었음을 암시했습니다.
시베리아 및 북동부 유럽 인구
집단의 데이터를 사용하여 분석한 결과, 우리는 이전에 보고된 논문 결과를 사실을 재확인하였고, 그리고 새로 현대와 고대 유라시아 인들 사이에
중요한 새로운 연결 고리를 발견 하였습니다. 우리는 동부 시베리아 인구가 그보다 오래된 다른 동아시아
인구에서 뚜렷이 분리된 하위 계급으로 형성했다는 것을 알게 되었고, 이 동부 시베리아는 ~8.8-11.2 kya에 동 아시아 그룹에서 분리된
것이다. 우리는 또한 시베리아 수렵
채집인과 동유럽 수렵 채집인인, 사마라(Samara),
카렐리야(Karelia), 헝가리(Hunggary), 스웨덴(Sweden)들이
(6.6-8 kya에서) 서로 혼혈 되었음을 확인했다. 또한 (4.7-5.3 kya)년에는 유럽인들의 조상이 되는 Yamnaya 사람들이 시베리아 수렵 채집인과 혼혈이
되었다. 그리고 현대 북동 유럽인들도 이때 혼혈이 되었다. 이는 시베리아 인들이 얼마나, 고대에도 그렇지만,
현대에도 유럽에 많은 유전적 기여를 했다는 것을 의미한다.(필자 멘트: 시베리안 북 유라시아인(=ANE)이 결국은 현대 유럽인들의 조상이라는
말이다.)
우리는 동아시아인과 시베리아인
사이에도 38%에 이르는 괄목할만한 유전적 영향을 우스트
이쉼 인들에게 주고 있다는 것 보게 된다. D-statistic은 또한 동아시아와 동 시베리아인들에게 현재 유럽보다 보다
강한 강한 친연성을
보인다. ( 그림
5 C; 보충 그림들 S13C,
S15 ). 동 아시아인과 동 시베리아인과 Ust'-Ishim의
친연성을 가리키는 또 다른 증거는 Y 염색체 haplogroup 분석에서도 제공되었다 ( 그림4A; Supplemental
Fig. S12). 우리는 Ust'-Ishim의 Y 염색체
haplogroup이 NO 계보(clade)에 속해 있고, NO 계보는 N의 조상으로, haplogroup N은 특히 시베리아와 북동부 유럽에서는 흔히 보이는 공통
유전자 이다. 또한 NO부터 아시아에 가장 흔한 haplogroup O가 분기
된다. 이러한 새로운 관계는 Mansi, Khanty, Nenets, Evens, Buryat
및 기타 시베리아 인을 포함한 서부 및 동부 시베리아 게놈을 포함하여 연계해 분석함으로써 그 관계가 크게
드러났습니다.그러므로 여기에 기술된 새로운 발견은 원래의
Ust'-Ishim 연구에서 관찰된 패턴의 보다 정확한 해석을 제공한다 ( Fu et al.,
2014 ).
(필자 멘트:
4,7만년의 우스트이쉼인들이 동아시아와 38% 유전적 공유하고 있다는 것은, 우스트이쉼인 들이 순다
대륙을 출발하여 동 아시아를 거쳐 우랄 툰드라에 진입했다는 걸 의미한다. )
특히 놀라운 발견은
2.4만년전의 시베리아 말타 소년, 아메리카 원주민, 서부 시베리아인 만시, 한티, 네네츠 사이의 유전적 연결
고리였습니다. 이 발견은 공통적인
'ANE(=고대 북 유라시아인, Ancient North
Eurasian)' 조상으로 인해 서부 시베리아인과 아메리카 인디언 사이의 강력한 유전적 관계를
입증합니다. 우리는 서부 시베리아
만시와 한티 계통의 57%가 '바이칼 말타 부렛'의 MA-1과 '아폰토바 고라' AG-2 고대인으로 대표되는 ANE
혈통과 연결되 있다고 추정한다. 다른 중요한 발견은 서부 시베리아 조상의 나머지 43 %가
동부 시베리아인과 관련이 있었기 때문에, 동부 시베리아 집단과 에벤키와 Evens 및 서부 시베리아인들의 공통 조상과 가장 밀접한 관련이
있음을 보여 준다 ( 그림
2). 이것은 현대 동북 유럽인들 간의 조상에 관련된 맥락에서
특히 흥미롭습니다. Lazaridis et al. (2014)(Mordovians), 핀란드 (Finns), 러시아
인, 사미 (Saami), 추바시 (Chuvash)와 같은 경우, ANE(=고대 유럽 북부 유라시아인 (MA-1로 대표)), WHG(=서유럽
수렵 채집인 (Loschbour 대표, WHG, West Hunter Gather)), 초기 NF(=유럽 신석기 농부(슈투트가르트 대표)) 으로 3 갈래로 조상이 형성 된다지만, 그러나 이
세가지 동유럽만 조상 간의
혼혈만으로는 그들의 형성이 설명되지는 않습니다. 이러한 점은 다른 서 유럽지역과는 달리, 동유럽인들은 동 아시아인들과 크게 연결되어 그렇다고만
알려졌었습니다. 그러나 Mezen
러시아인, Komi, Karelians 및 Veps와 같은 동북 유럽 인구의 유전적 관계에 대한 우리의 분석은, 현대 유럽인들 사이에서도 설명되지 않는 조상 구성 요소가 서부 및 동부 시베리아 인구와의
친연성에 기원한 것으로 보인 것으로 나타났습니다.
이와 더불어, 우리의 이와 같은 새로운 발견이 보여 주는 것은, 또한 서부 시베리아인 만시 (Mansi)와 한티 (Khanty)와
'고대 동유럽 인구 (=Yamnaya와 PWC 인)'가 상당량의 ANE와 조상을 공유했다는 것을 보여
주었다. 동부
시베리아인들과 고대 동유럽인들의 유전적 친화성은 상대적으로 약하다 ( 그림
5 E; 보충 그림들 S21,
S24 ). 그러므로 우리는 북동부 유럽인들에 대한 서부 시베리아 섞임이 Yamnaya 문화가 시작되기 전에
(5.3-4.7 kya) 일어 났다고 본다. Mansi와의 혼합이 북동부 유럽의 수렵 채집인들 사이에서 6.6-8 kya
(카렐리아 수렵 채집인 , 사마라 수렵 채집인 및 Mtala수렵 채집인 및 헝가리 감바 수렵 채집인) ( 보충 그림
S21F-Q) 그
Yamnaya 사람들이 Pre-Yamnaya 사람들은 CHG(Caucus Hunter
gather)와 상당 부분 유전자 공유를 하지만, 만시(Mansi)는 Yamnaya 사람들과 달리 후기
CHG와는 조상을 공유하지 않는데 ( Supplemental
Fig. S25 ), 이것이 의미하는 바는. Mansi가 Yamnaya
사람들의 후손이 아님을 시사한다. 즉 만시가 얌나야의 조상이기 때문에, 따라서 북동부 유럽으로의 시베리아
섞임은 6.6 kya
이전에 시베리아인과 북동부 유럽인
(7.1-4.9 kya) 사이에서 Y 염색체 haplogroup N1c1의
확장과 동시에 시작되었을 가능성이 크다.
haplogroup N은 동아시아에서 유래했을 가능성이 높고
( Shi et al.,
2013 ) 현재 시베리아 인구 중 가장 높은 빈도를
보이고 있기
때문에 동유럽인들 사이에
시베리아에서 북동부 유럽으로의 고대 유전자 흐름이 반영될 가능성이 높습니다.
우리의 결과는 또한
Uralic 지역의 원주민 집단의 기원에 관한 인류학적 논쟁에 대한 통찰력을 제공합니다. 우랄(Uralic) 사람들은 유럽인과 아시아인 사이의 오랜
혼합에 기인한 혼합 그룹으로 여겨졌다. 대안적인 이론은 우랄 사람들이 유럽과 아시아 계보
( Bunak
1956 ) 와는 다른 유전적 혈통을 형성했다는 가설을
세웠다. 이 아이디어는 특이한 인류학적 복합체가 관찰된 북부 우랄릭
지역의 인종 집단에 대한 광범위한 인류학 연구를 기반으로 했다 ( Cheboksarov
and Trofimova 1941 ; Bunak
1956 , 1965). 현재까지 이 가설은 거부되거나 확인되지
않았습니다. 유럽, 동아시아,
시베리아 ( 그림
8 ) 의 지난 5 만년 동안의 유전자 흐름
모델은 서 시베리아 사람들이 ANE
사람들과 관련이 있을 수 있는 고대 혈통과 뚜렸d하게 관련되어 있었기 때문에 Bunak's hypothesis을 뒷받침한다. 동시에, 우리 모델은 비교적
근자인 7~9 kya 고대 서부 시베리아인들 (서부
시베리아와 접촉한 사람들)과의
혼혈을 포함하는 보다 복잡한 시나리오를 제안합니다.
요약하면,
우리의 연구는 지난 45,000 년 동안 시베리아, 유럽 및 동아시아 인구의 고대 유전적 연결 고리를
확립했습니다.우리는
시베리아와 동유럽에서 수집된 개체군의 특징적인 게놈 관계가 여전히 희박하다는 것을 알고 있지만 우리 연구에서 밝혀진 새로운 유전적 연결은
시베리아와 동유럽 인구에 대한 유전 및 언어 연구에 대한 기초를 제공할 것으로 기대합니다.
필자의
멘트:
필자는 산스크리트어를 연구하다가 왜 이러 언어의
유사성이 성립하는 지를 연구하다가 이런 유전학을 공부하게 되었다.
필자가 주장하는 것은 아래와
같다.
-Oppenheimer(2012)의 초기
이동(흑백)위에 필자가 붉은 색으로 추가하였다.
-최대빙하기를 기준으로 하면, 강수량이 현재에 비해 1/10~1/20으로 매우 적어,
내륙은 거의 사막이다. 동아시아를 제외한 유라시아 대륙은 거의 사막이기 때문에, 빙하기 때 인구 이동은 내륙에서 이동은 많이 제한되었다. 따라서 out Of Africa에 이은 1차 해안가 이동과 순다 대륙에서 한반도에 이르는 태평양 연안 뻘 밭이 주요한 확산 루트이고, 그 다음
단계로 물이 풍부하고 거대 동물 떼가 흔한 툰드라 스텝이 2차로 매우 중요해 진다.
따라서 크게 보면 순다
대륙에서 반 시계 방향으로 중국 태평양 해안가를 통해 북상하고, 툰드라 스텝을 통해 확산한 것이 주요한 초기 이동
경로이다. 이러한 필자의 가설은 이 논문에서 분명히 보여주는 고인골 유전자 분석 결과로 뒷 받침하여 준다. 이 논문의 결과에 따르면, 논문의 '그림8'과는 달리 다음과 같이 그려져야 할 것이다. 논문의 '그림 8'은 많은 오해의
소지가 있는 그림이다. 다음 그림에서 필자가 추가하여 그린 것은 빨간색이다. 이 논문의 말타와 관련된 57%도 순다 대륙의 K계열에서 분기하여
유럽에서 성장한 것이기 때문에 '그림8'의 그림은 정확치가 않다.
순다 대륙에서 우랄 평원의 우스트이쉼의 진입은 인도네시아에서 남쪽으로 시계 방향으로,
인도를 통해 중앙아시아를 거쳐 진입하는 것도 하나의 가능성이 있는 루트이데, 이는 시계 방향이다. 또 다른 하나는 중국 동해안을 거쳐, 한반도를
거쳐 툰드라로 진입하는 반 시계 방향이다. 지금의 일반론적인 정설은 시계 방향이다(아래 그림 검은색 참조 ).
그러나 필자의 언어학에 기반한 결론은 반시계 방향으로 진행되는 것이다. 이는 현재의
통용되는 일반 정설과는 다른 견해이지만, 필자는 2013년부터 언어 분석에 기초하여 처음부터 주장한 내용이 이것이다. 여기 이 논문은 이런
필자의 가설들을 드디어 하나 씩 증명해 주기 시작한다.
즉 빨간색 점 선은 시계
방향으로 남쪽에서 유럽으로 진입한 유전자인데, 이 루트도 분명히 있긴 있겠지만, 생각보다 이 부분이 차지하는 비중은 크지가 않다. 따라서
Main Stream은 인도네시아에서부터 툰드라를 통해 유럽에 진입한 것이 주력이다. 물론 이후에는 동서로의 이동이 여러 차례 발생했을
것이지만, 큰 틀에서는 동에서 서로 진행된 것이다.
이런 필자의 견해와 이
논문의 해석의 차이를 비교해 보라. 물론 지금 남방의 검은 피부가 아닌 사람들은, 모두 북쪽에서 남하하여 남쪽으로 역 확산한 사람들이다. 다시
말하면, 툰드라에서 남방으로 역 확산도 광범위하게 일어 났다. 예를들면 인도의 타밀어 화자들은 이런 경로는 형성되었다고 보면 될
것이다.
물론 이 논문에서 지적한
것처럼, 빙하기 이후에 1만년 후에도 Y-염색체 N으로 대표되는 본격적인 이동도 포함된다. 이를 보면, 아주 고대에 툰드라를 통한 인류 이동도
그랬지만, 그 이후에도 끊임없이 동에서 서로 유전자 흐름이 있어 왔다는 것은 이처럼 분명한
사실이다.
참고로 중화문명의 원조라
여겨지는 중원 지역의 앙소 문화는 바로 이러 북방 노마드인들이 세운 무명으로, 현재의 중국인과는 별 상관이 없다. 이런 경향은 회수 북방의
중국 지역에서 공통적으로 나타나는 현상으로, 앙소문화를 뒤따르는 용산 문화 등에도 나타나는 현상이다. 이보다 더 북방인 홍산 문명은 말할 것도
없다. BC1,600년 이후에나 간헐적으로 등장하는데, 이들은 북방 노마드 문명이다. 이들 'N'형은 동유럽 형성에도 많은 유전적 영향을
끼쳤다.
따라서 '중화'라는 것은
그냥 정치적으로 형성된 개념일 뿐으로 과학적 근거가 없다는 것이다.
다음 그림은 필자가 2013년에 그린 그림인데, 지금
봐도 크게 무리가 없는 그림이어서 인용한다.
우리는 시베리아와 동유럽에서 온 14 개의 인종 그룹을
대표하는 28 명의 개인을 심층적으로 분류했습니다. 모집단 데이터의 질을 극대화하기 위해 우리는 전통적으로
해당 원주민에 의해 점유된 지리적 위치와 최소한 3세대 동안 유전자 혼합이 없는 개인에서 유익한 DNA 표본을 얻었다. 우리의 샘플은 시베리아와 동유럽의 주요 인구를 대상으로
하며,이 지역의 주요 언어 그룹을 대표합니다 ( 표
1 ; 그림
1 ).
혈액
샘플은 러시아 과학 아카데미 분자 유전학 연구소 (Institute of Molecular Genetics)의 IRB 승인 승인하에 정보에 입각
한 동의하에 수집되었습니다. 남부
캘리포니아 대학 (University of Southern California)의 IRB 승인 (HS-13-00594)에 따라 샘플을 더 이상
확인하지 않고 분석했습니다. DNA는 프로 테이나 제 K 처리 및 페놀- 클로로포름
추출 ( Milligan
1998 )을 사용하는 표준 기술에 따라 말초 백혈구로부터
분리하였다. Illumina 플랫폼 (HiSeq 및 X10)에서 모든
DNA 샘플의 시퀀싱을 평균 시퀀스 커버리지 깊이가 38x 인 최소 30x (고유 매핑된 읽기 기반)의 깊이로
시퀀싱했습니다. 모든 게놈은 동일한
계산 도구 세트 (Novoalign과 GATK)를 사용하여 분석되었습니다 ( McKenna 외.
2010) 및
필터링 기준. 모든 분석에서
SNP를 호출하기 위해 mapQ 점수가 60 이상인 정렬을 사용했습니다. 이들 샘플의 Ti / Tv
비율은 보충 표
S5에 나열되어있다. 뿐만 아니라 Mansi와 Khanty 두 서부 시베리아
인구의 많은 게놈 데이터 세트가 없었기 때문에 고밀도 SNP 마이크로 어레이를 사용하여 이 두 개체군 (각각 45 명과 37 명)의 표본을
유전자형으로 분류하여 인구 관계 및 표본 품질을 확인했습니다. 전체 게놈 시퀀싱 데이터.
세계의
다른 개체군과 비교하여 시베리아와 동유럽 개체군의 유전적 관계를 조사하기 위해 18 개 개체군을 대표하는 32 개의 높은 범위의 최신
게놈으로부터 공개적으로 이용 가능한 원시 염기 서열 데이터를 통합했다 ( Supplemental
Table S2 ; Wong et al.
2013 , 2014 ; Zhou . 등
2013 ; 정 등
2014. ; . Prufer 등
2014 ), 고대의 게놈
( 보충 표
S3 )및 변이 로 2개
호미닌의 게놈, 즉 네안데르탈인 ( Prufer 등
2014 )과 데니소바인
( 마이어 등 2012
년)를
분석합니다. 다른 시퀀싱 플랫폼,
다양한 읽기 매핑 도구, 다양한 SNP 도구 및 다운 스트림 변형 필터의 사용으로 인한 잠재적인 편향을 최소화하고자
했습니다. 따라서 Denisova,
Neanderthal 및 genotyped 샘플을 제외한 모든 게놈은 이 연구에서 다른 게놈과 동일한 도구, 매개 변수 및 필터링 기준을
사용하여 초기 시퀀싱 읽기부터 재분석되었습니다. 원래 연구와의 유전자형 품질 및
일치도는 보충 표
S4에 나와있다 .
TreeMix
우리는
상 염색체 TreeMix ( Pickrell and
Pritchard 2012 ) 게놈 전체 상 염색체 SNP 30,090,159
세트를 사용하여 그래프를 작성했습니다. TreeMix는 분기 트리 형태의 인구 통계 시나리오를
모델링하여 과거와 과거의 숨겨진 인구 통계학적 이벤트에 대한 통찰력을 제공하기 위해 개인과 인구 간의 혼혈 이벤트를 추론 할 수
있습니다.
D- 통계 테스트
TreeMix에 의해 추론 된 혼합 이벤트에 대한 통계적
지원과 관심 대상 집단 간의 관계를 조사하기 위해 ADMIXTOOLS를 사용하여 D (P 1 , P 2 , P 3 , O) 형태의 D 통계 테스트를 수행 했습니다 ( Patterson et
al. 2012 ) 릴리스 1.1. ADMIXTOOLS는 다음 표기법을
사용합니다. D (W, X, Y,
Z) . 우리의 파라미터는 다음과 같은 방법으로 ADMIXTOOLS
파라미터에 대응 : O =W, P (1)=Z, P 2=Y, P (3)=의X . 이 테스트에서 귀무 가설은 트리 토폴로지
((( P 1 , P2 ), P 3 ), O)가 정확하고 P 3 과 P 1 또는 P 2 사이의 유전자 이동이나 이들과 관련된 개체군이
없다. D-
통계 테스트는 데이터가 가설과 일치하지 않는지 평가하는 데 사용할 수 있습니다.
MSMC 분석
집단 간 분기
시간을 추정하기 위해 MSMC ( Schiffels
and Durbin 2014 )를 4 개 또는 8 개 haplotype 중 하나에서
실행했습니다. 즉, 두 개체군
각각으로부터의 두 개의 Haplogroup 또는 두 개체군 각각으로부터의 네 가지 haplo를 포함한다. 5 개의 샘플 (프랑스인, 러시아인, Mansi,
Evenki 및 Han)을 모집단으로 사용하고 다른 집단과의 분기 시간을 계산했습니다. 분석된 각 모집단에 대해, 우리는 5개의 참고 인구 집단
각각에 대한 평균 분화된 시간 추정치를 보여 줍니다. 돌연변이 속도와 세대 당 년수에 대해 Schiffels and Durbin
(2014)에서와 같이 연간 bp 당 1.25 x 10 -8 , 30 yr를 사용했습니다.
Y 염색체와
미토콘드리아 DNA에 대한 SNV와 haplogroup의 분석은 보충
자료 와 보조 수치 S10 ~
S12 에서 찾을
수 있습니다 .
The 1000 Genomes Project Consortium. 2012. An integrated
map of genetic variation from 1,092 human genomes. Nature491:
56–65. [PMC free
article][PubMed]
Allentoft ME, Sikora M, Sj?gren KG, Rasmussen S, Rasmussen
M, Stenderup J, Damgaard PB, Schroeder H, Ahlstr?m T, Vinner L, et al.
2015. Population genomics of Bronze Age Eurasia. Nature522:
167–172. [PubMed]
Bunak VV. 1956. Human races and
ways of their formation. Sov
Ethnogr1: 86–105 (in Russian).
Bunak VV. 1965. Problems of the
genesis of races. In The origin and
ethnic history of Russian people (ed. Bunak VV), pp. 174–190. Nauka,
Moscow (in Russian).
Cheboksarov NN, Trofimova TA. 1941. Anthropological
survey of Mansi. Short Reports of Institute of History of Material
Culture9: 28–36.
Der Sarkissian C, Balanovsky O, Brandt G, Khartanovich V,
Buzhilova A, Koshel S, Zaporozhchenko V, Gronenborn D, Moiseyev V, Kolpakov E,
et al. 2013. Ancient DNA reveals prehistoric gene-flow from Siberia in the
complex human population history of North East Europe. PLoS
Genet9: e1003296. [PMC free
article][PubMed]
Durand EY, Patterson N, Reich D, Slatkin M. 2011. Testing for
ancient admixture between closely related populations. Mol Biol
Evol28: 2239–2252. [PMC free
article][PubMed]
Erdeniev UE. 1985. Kalmyks. Nauka, Moscow.
Fedorova SA, Reidla M, Metspalu E, Metspalu M, Rootsi S,
Tambets K, Trofimova N, Zhadanov SI, Hooshiar Kashani B, Olivieri A, et al.
2013. Autosomal and uniparental portraits of the native populations of
Sakha (Yakutia): implications for the peopling of Northeast
Eurasia. BMC Evol Biol13:
127. [PMC free
article][PubMed]
Fu Q, Li H, Moorjani P, Jay F, Slepchenko SM, Bondarev AA,
Johnson PL, Aximu-Petri A, Prufer K, de Filippo C, et al. 2014. Genome sequence
of a 45,000-year-old modern human from western Siberia. Nature514:
445–449. [PMC free
article][PubMed]
Gamba C, Jones ER, Teasdale MD, McLaughlin RL,
Gonzalez-Fortes G, Mattiangeli V, Dombor?czki L, Ko˝v?ri I, Pap I, Anders A, et
al. 2014. Genome flux and stasis in a five millennium transect of European
prehistory. Nat Commun5:
5257. [PMC free
article][PubMed]
Haak W, Lazaridis I, Patterson N, Rohland N, Mallick S,
Llamas B, Brandt G, Nordenfelt S, Harney E, Stewardson K, et al. 2015. Massive
migration from the steppe was a source for Indo-European languages in
Europe. Nature522:
207–211. [PMC free
article][PubMed]
Hublin JJ. 2009. Out of Africa:
modern human origins special feature: the origin of Neandertals. Proc Natl Acad
Sci106: 16022–16027. [PMC free
article][PubMed]
Hublin JJ. 2012. The earliest
modern human colonization of Europe. Proc Natl Acad
Sci109: 13471–13472. [PMC free
article][PubMed]
The International HapMap 3 Consortium. 2010. Integrating
common and rare genetic variation in diverse human populations. Nature467:
52–58. [PMC free
article][PubMed]
International Society of Genetic Genealogy. 2014. Y-DNA Haplogroup
Tree 2014, Version: 9.76, Date: 29 August 2014, http://www.isogg.org/tree/.
Jeong C, Alkorta-Aranburu G, Basnyat B, Neupane M, Witonsky
DB, Pritchard JK, Beall CM, Di Rienzo A. 2014. Admixture
facilitates genetic adaptations to high altitude in Tibet. Nat
Commun5: 3281. [PMC free
article][PubMed]
Jones ER, Gonzalez-Fortes G, Connell S, Siska V, Eriksson
A, Martiniano R, McLaughlin RL, Gallego Llorente M, Cassidy LM, Gamba C, et al.
2015. Upper Palaeolithic genomes reveal deep roots of modern
Eurasians. Nat Commun6:
8912. [PMC free
article][PubMed]
Khrunin AV, Khokhrin DV, Filippova IN, Esko T, Nelis M,
Bebyakova NA, Bolotova NL, Klovins J, Nikitina-Zake L, Rehnstrom K, et al.
2013. A
genome-wide analysis of populations from European Russia reveals a new pole of
genetic diversity in northern Europe. PLoS
one8: e58552. [PMC free
article][PubMed]
Lappalainen T, Laitinen V, Salmela E, Andersen P, Huoponen
K, Savontaus ML, Lahermo P. 2008.Migration waves
to the Baltic Sea region. Ann Hum
Genet72(Pt 3): 337–348. [PubMed]
Lazaridis I, Patterson N, Mittnik A, Renaud G, Mallick S,
Kirsanow K, Sudmant PH, Schraiber JG, Castellano S, Lipson M, et al. 2014. Ancient human
genomes suggest three ancestral populations for present-day
Europeans. Nature513:
409–413. [PMC free
article][PubMed]
Li JZ, Absher DM, Tang H, Southwick AM, Casto AM,
Ramachandran S, Cann HM, Barsh GS, Feldman M, Cavalli-Sforza LL, et al.
2008. Worldwide human relationships inferred from genome-wide patterns
of variation. Science319:
1100–1104. [PubMed]
Malyarchuk B, Derenko M, Grzybowski T, Lunkina A, Czarny J,
Rychkov S, Morozova I, Denisova G, Miscicka-Sliwka D. 2004. Differentiation
of mitochondrial DNA and Y chromosomes in Russian populations. Hum
Biol76: 877–900. [PubMed]
McKenna A, Hanna M, Banks E, Sivachenko A, Cibulskis K,
Kernytsky A, Garimella K, Altshuler D, Gabriel S, Daly M, et al. 2010. The Genome
Analysis Toolkit: a MapReduce framework for analyzing next-generation DNA
sequencing data. Genome Res20:
1297–1303. [PMC free
article][PubMed]
Meyer M, Kircher M, Gansauge MT, Li H, Racimo F, Mallick S,
Schraiber JG, Jay F, Prufer K, de Filippo C, et al. 2012. A high-coverage
genome sequence from an archaic Denisovan individual. Science338:
222–226. [PMC free
article][PubMed]
Milligan BG. 1998. Total DNA
isolation. In Molecular genetic analysis of populations (ed. Hoelzel
AR), pp. 29–60. Oxford University Press, Oxford.
Mirabal S, Regueiro M, Cadenas AM, Cavalli-Sforza LL,
Underhill PA, Verbenko DA, Limborska SA, Herrera RJ. 2009. Y-Chromosome
distribution within the geo-linguistic landscape of northwestern
Russia. Eur J Hum Genet17:
1260–1273. [PMC free
article][PubMed]
Patterson N, Price AL, Reich D. 2006. Population
structure and eigenanalysis. PLoS
Genet2: e190. [PMC free
article][PubMed]
Patterson N, Moorjani P, Luo Y, Mallick S, Rohland N, Zhan
Y, Genschoreck T, Webster T, Reich D. 2012.Ancient
admixture in human history. Genetics192:
1065–1093. [PMC free
article][PubMed]
Peoples of Russia. 1994. Encyclopaedia (ed. Tishkov VA), 479 pages. Bol'shaya
Sovetskaya Entsyklopediya, Moscow (in Russian).
Pickrell JK, Pritchard JK. 2012. Inference of
population splits and mixtures from genome-wide allele frequency
data. PLoS Genet8:
e1002967. [PMC free
article][PubMed]
Poznik GD, Henn BM, Yee MC, Sliwerska E, Euskirchen GM, Lin
AA, Snyder M, Quintana-Murci L, Kidd JM, Underhill PA, et al. 2013. Sequencing Y
chromosomes resolves discrepancy in time to common ancestor of males versus
females. Science341:
562–565. [PMC free
article][PubMed]
Price AL, Patterson NJ, Plenge RM, Weinblatt ME, Shadick
NA, Reich D. 2006. Principal components analysis corrects for stratification in
genome-wide association studies. Nat
Genet38: 904–909.[PubMed]
Prufer K, Racimo F, Patterson N, Jay F, Sankararaman S,
Sawyer S, Heinze A, Renaud G, Sudmant PH, de Filippo C, et al. 2014. The complete
genome sequence of a Neanderthal from the Altai Mountains. Nature505:
43–49. [PMC free
article][PubMed]
Purcell S, Neale B, Todd-Brown K, Thomas L, Ferreira MA,
Bender D, Maller J, Sklar P, de Bakker PI, Daly MJ, et al. 2007. PLINK: a tool
set for whole-genome association and population-based linkage
analyses. Am J Hum Genet81:
559–575. [PMC free
article][PubMed]
Raghavan M, Skoglund P, Graf KE, Metspalu M, Albrechtsen A,
Moltke I, Rasmussen S, Stafford TW Jr, Orlando L, Metspalu E, et al. 2014. Upper
Palaeolithic Siberian genome reveals dual ancestry of Native
Americans. Nature505:
87–91. [PMC free
article][PubMed]
Rasmussen M, Li Y, Lindgreen S, Pedersen JS, Albrechtsen A,
Moltke I, Metspalu M, Metspalu E, Kivisild T, Gupta R, et al. 2010. Ancient human
genome sequence of an extinct Palaeo-Eskimo. Nature463:
757–762. [PMC free
article][PubMed]
Schiffels S, Durbin R. 2014. Inferring human
population size and separation history from multiple genome
sequences. Nat Genet46:
919–925. [PMC free
article][PubMed]
Shi H, Qi X, Zhong H, Peng Y, Zhang X, Ma RZ, Su B.
2013. Genetic evidence of an East Asian origin and paleolithic
northward migration of Y-chromosome haplogroup N. PLoS
one8: e66102. [PMC free
article][PubMed]
Skoglund P, Malmstrom H, Raghavan M, Stora J, Hall P,
Willerslev E, Gilbert MT, Gotherstrom A, Jakobsson M. 2012. Origins and
genetic legacy of Neolithic farmers and hunter-gatherers in Europe. Science336:
466–469. [PubMed]
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar
S. 2011. MEGA5: molecular evolutionary genetics analysis using maximum
likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol
Evol28: 2731–2739. [PMC free
article][PubMed]
Waters MR, Forman SL, Pierson JM. 1997. Diring Yuriakh:
a lower paleolithic site in central Siberia. Science275:
1281–1284. [PubMed]
Wong LP, ong RT, Poh WT, Liu X, Chen P, Li R, Lam KK,
Pillai NE, Sim KS, Xu H, et al. 2013. Deep
whole-genome sequencing of 100 southeast Asian Malays. Am J Hum
Genet92: 52–66. [PMC free
article][PubMed]
Wong LP, Lai JK, Saw WY, ong RT, Cheng AY, Pillai NE, Liu
X, Xu W, Chen P, Foo JN, et al. 2014.Insights into
the genetic structure and diversity of 38 South Asian Indians from deep
whole-genome sequencing. PLoS
Genet10: e1004377. [PMC free
article][PubMed]
Yunusbayev B, Metspalu M, Jarve M, Kutuev I, Rootsi S,
Metspalu E, Behar DM, Varendi K, Sahakyan H, Khusainova R, et al. 2012. The Caucasus as
an asymmetric semipermeable barrier to ancient human migrations. Mol Biol
Evol29: 359–365. [PubMed]
Zhou D, Udpa N, Ronen R, Stobdan T, Liang J, Appenzeller O,
Zhao HW, Yin Y, Du Y, Guo L, et al. 2013.Whole-genome
sequencing uncovers the genetic basis of chronic mountain sickness in Andean
highlanders. Am J Hum Genet93:
452–462. [PMC free
article][PubMed]