<P>사실은..,&nbsp; 실험도 애저녁에 끝나 예상된 결과를 가지고 있지만...</P>
<P>남들 하는대로 '계획서'로 ....&nbsp; </P>
<P>&nbsp;</P>
<P>유심히 봐 주실분이 몇이나 되실런지...ㅋㅋㅋㅋ</P>
<P>&nbsp;</P>
<P>&nbsp;</P>
<P>&nbsp;</P>
<P>&nbsp;</P>
<P>&nbsp;</P>
<P>&nbsp;</P>
<P><B style="FONT-WEIGHT: normal" id=docs-internal-guid-42b1c035-9c95-9e78-7264-bb9ec7ac7730>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 19px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp;&nbsp; 석사 학위 청구 논문 계획서</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P><BR><BR>&nbsp;</P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 28px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">구속 스트레스 후 자발적인 운동이 &nbsp;&nbsp;</SPAN></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 28px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">스트레스 호르몬 및 세포사멸에</SPAN></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 28px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">미치는 영향</SPAN></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P><BR><BR><BR>&nbsp;</P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><BR>&nbsp;</P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: #ffffff; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 21px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: bold; TEXT-DECORATION: none">목 차</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: bold; TEXT-DECORATION: none">Ⅰ. 서론···································································································1</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">1. 연구의 필요성·······················································································1</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">2. 연구의 목적························································································0</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">3. 연구의 가설························································································0</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: bold; TEXT-DECORATION: none">Ⅱ. 이론적 배경·······················································································0</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">1. 스킨스쿠버 다이빙··············································································0</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">2. 구속스트레스·····················································································0</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">3. 운동과 스트레스·················································································0</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">4. 자발적 운동과 스트레스 ·····································································0</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt; MARGIN-RIGHT: 2pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">5. 운동과 세포사멸················································································0</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: bold; TEXT-DECORATION: none">Ⅲ. 연구방법··························································································0</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">1. 실험동물 및 사육방법··········································································0</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">2. 식이 및 섭취방법················································································0</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">3. 구속스트레스 방법··············································································0</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">4. 운동방법···························································································0</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">5. 시료채취···························································································0</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">6. 분석항목 및 방법················································································0</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">1) 스트레스 호르몬 분석··········································································0</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">2) 단백질 발현 분석················································································0</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">7. 자료 처리 방법···················································································0</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 17px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">참고문헌······························································································0</SPAN></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P><BR><BR><BR><BR><BR>&nbsp;</P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: -35pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 19px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: bold; TEXT-DECORATION: none">I. 서론</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">1. 연구의 필요성</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">스트레스 본질의 개념은 “물체에 가해지는 물리적 힘(단위면적당 하중)”을 의미하였지만 사회와 시대가 급속도로 변화함에 따라 의학적인 개념으로 응용되어지고 있다. 즉, 의학적으로 스트레스란 신체 또는 심리적으로 평형상태를 이루지 못하여 각종 질병 및 질환의 요인이 되며, 이로 인한 환자 수는 급증하고 있다. </SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">반복스럽고 혐오스러운 상황은 동물에게 만성적인 스트레스가 될 수 있으며, 시상하부-뇌하수체-부신 축(hypothalamic-pituitary adrenal axis)의 만성적인 활성에 의해 생리적 변인(체중, 혈액 생화학 변인)과 행동(감정적 반응, 인지반응, 정형화된 사고 등)의 변화가 나타날 수 있다(Laurence et al., 2014). 또한 단기간 스트레스는 동물에게 다양한 호르몬 변화의 시발점이 될 수 있다(Jameel et al., 2014). &nbsp;구속 스트레스를 부하하게 되면 노화 억제의 시발점이 되는 단백질인 Bcl2 mRNA가 유의하게 감소하며, 난소의 에스트라디올, 테스토스테론, IGF-I이 감소하는 반면에 코티졸과 프로게스테론은 유의하게 증가한다(Liang et al., 2014). 보다 다른 관점에서 본다면, 만성적인 스트레스는 섭취음식을 선택할 때, 탄수화물과 포화지방산을 선호하게 되는 것으로 나타나 체중이 증가하게 되는 원인이 되기도 한다(Robert et al., 2014). 구속스트레스는 키노몰루스 원숭이(Cynomolgus monkey)의 글루코스 농도 및 코티졸 농도를 증가시킨다(Shirasaki et al., 2013).</SPAN></P>
<P><BR><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">한편, 스쿠버 다이빙은 스트레스 호르몬인 혈중 코티졸 농도를 증가시킨다(</SPAN><A style="TEXT-DECORATION: none" href="http://www.ncbi.nlm.nih.gov/pubmed?term=Zarezadeh%20R%5BAuthor%5D&amp;cauthor=true&amp;cauthor_uid=25311329"><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Zarezadeh </SPAN></A><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">&amp; </SPAN><A style="TEXT-DECORATION: none" href="http://www.ncbi.nlm.nih.gov/pubmed?term=Azarbayjani%20MA%5BAuthor%5D&amp;cauthor=true&amp;cauthor_uid=25311329"><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Azarbayjani</SPAN></A><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">, 2014). 또한 Perovic et al. (2014)은 스쿠버 다이빙 시 수중에서의 신체활동뿐만 아니라 수중에서 전개되는 다양한 환경적 요인에 의해 활성산소 생산이 증가하게 되어 산화적 손상(oxidative damage)이 매우 증가한다고 하였다. 스쿠버 다이빙은 과산소(hyperoxia)와 과분압(hyperbaria)의 특징이 있으며, 활성산소종(reactive oxygen species: ROS)를 증가시켜 산화스트레스를 유발하기도 한다(Sureda et al., 2014). 또한, 스쿠버 다이빙은 산화스트레스를 유발하는 운동의 한 종류로서 신체활동에 의한 것뿐만 아니라 수중에서의 환경적 요인의 변화에 대해서도 산소유리기(free radicals: FR)의 발생을 증가시키기도 한다(Perovic et al., 2014). FR은 노화의 주된 원인 중 하나이다(Park et al., 2014). </SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">한편, 운동은 스트레스의 저항성을 개선하고 흥분을 억제시키며, 사람과 동물모델에서 우울증을 감소시키기도 한다(Hare et al., 2014). 암컷 쥐에게 반복적인 고립스트레스 후 장기간 트레드밀 달리기 운동훈련을 부하하였을 때 호르몬 농도가 개선되었다는 보고(Han et al., 2014)도 있다. 또한 뇌졸중 유발 이후 사지의 자발적인 움직임은 손상된 사지의 기능성 회복에 도움을 준다(Livinston-Thomas et al., 2014). Han et al.(2014)은 반복적인 고립스트레스를 부하하였을 때 장기간 운동이 스트레스를 해소시키며, 동시에 강제수영운동의 활동성을 개선하였다고 보고하였다. 구속 스트레스는 강제수영운동에 비하여 더 큰 스트레스가 주어지는 방법이다(Jameel et al., 2014).</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">만성적인 스트레스는 피부의 시상하부-뇌하수체-부신피질 축 요소와 멜라닌 생합성을 억제한다(Pang et al., 2014). 뿐만 아니라 Uribe et al.(2104)은 시상하부-뇌하수체 갑상선 축(hypothalamic-pituitary thyroid axis: HPT axis)이 구속스트레스, 가벼운 스트레스 혹은 식이제한에 의해 활성이 억제되었으나, 자발적 운동을 하였을 때는 관련이 없었다고 하였다. &nbsp;Castilla-Ortega et al. (2014)은 만성적인 스트레스에 이후 자발적인 운동이 해마 뉴런을 눈에 띄게 증가시키며 스트레스로 인한 ‘무엇을-언제-어디서’(what-when-where)와 관련된 기억력 손상을 예방한다고 하였다. 뿐만 아니라 만성적 구속스트레스에 의해 유발된 장관기능과 항균기능 저해가 가벼운 활동에 의해 억제된다는 보고도 있다(Luo et al., 2014). &nbsp;그러나 구속스트레스 이후에 강제운동을 부하하게 되면 스트레스에 의한 기억력 손상에 대한 운동의 효과가 나타나지 않는다는 연구결과도 있다(Radahmadi et al., 2013). 이와 유사하게 자발적 운동에 비하여 비자발적 운동을 부하하였을 때 실험 쥐의 불안 수준이 증가되며, 해마의 신경생합성에 작용하는 VEGF 및 BDNF의 수준이 감소하는 것으로 보고되었다(Uysal et al., 2014). </SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Heat shock protein 72 (HSP 72)는 운동에 의해 발현이 증가하게 되는 대표적인 스트레스 단백질로 활용되는 마커이다(Chang et al., 2014). 운동에 의해 증가하는 Mn isoform of superoxide dismutase (Mn-SOD)의 발현량 증대는 노화와 관련된 단백질인 Bax와 caspase-3 발현을 억제하여 노화 쥐 심장의 세포사멸을 감소시킨다(Kwak et al., 2014). 그러나 운동 강도가 높은 경우에는 반대의 결과를 나타낼 수 있다. Ding et al. (2014)은 고강도 운동이 과도한 세포사멸을 유도한다고 보고한 바 있다. 또한, 고강도 운동은 백혈구의 세포사멸을 유발하기도 한다(</SPAN><A style="TEXT-DECORATION: none" href="http://www.ncbi.nlm.nih.gov/pubmed?term=Kr%C3%BCger%20K%5BAuthor%5D&amp;cauthor=true&amp;cauthor_uid=25063943"><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Kr&uuml;ger</SPAN></A><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none"> &amp; </SPAN><A style="TEXT-DECORATION: none" href="http://www.ncbi.nlm.nih.gov/pubmed?term=Mooren%20FC%5BAuthor%5D&amp;cauthor=true&amp;cauthor_uid=25063943"><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Mooren</SPAN></A><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">, 2014).</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">한편, 중등도 강도의 트레드밀 운동 또는 자발적 운동은 세포사멸을 억제(Chung et al., 2014; Mokhtari-Zaer et al., 2014)하는 것으로 나타나며, 당뇨 유발 어미 쥐에서 태어난 생후 4주령된 새끼 쥐에게 트레드밀 운동은 세포사멸을 억제하여 단기기억 손상을 억제한다는 보고(Kim et al., 2014)도 있다. 또한 강제수영운동 또는 구속스트레스를 부하하였을 때 혈중 코티졸 농도가 동일하게 상승하였다는 연구결과도 보고되었다(Jameel et al., 2014). 이와 유사하게 자발적 운동은 약물 중독 이후의 금단증상 및 스트레스를 억제하지만, 강제운동에서는 그렇지 않다는 보고도 있다(Ogbonmwan et al., 2014). 마우스를 훈련시키는 방법 중 가장 일반적인 것은 전기충격이 이루어지는 상태에서 트레드밀 달리기를 이용한 지구성 운동방법으로서 마우스는 전기자극을 회피하기 위하여 트레드밀 벨트 위를 달리게 된다. 그러나 이때 주어지는 스트레스는 지구력 운동량과 운동 후 혈중 호르몬 및 사이토카인 수준에 변화를 주곤 한다(Conner et al., 2014). 이와 같은 보고는 강제운동에 비하여 자발적인 운동을 부하하였을 때 나타나는 결과가 보다 신뢰성 있는 결과를 제공할 수 있을 것이다. 따라서 구속스트레스 이후에 강제운동 보다는 자발적인 운동참여가 효과적일 것이라 판단된다. </SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">&nbsp;</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">2. 연구의 목적</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">본 연구에서는 스킨스쿠버 다이빙 상황과 유사한 구속스트레스를 인위적으로 유발한 후 자발적인 운동이 스트레스 호르몬 과 노화관련 단백질 발현의 상호기전을 규명하고 스트레스 상황 후 자발적인 운동의 효과 및 생리적 현상에 대하여 입증하고자 한다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">3. 연구의 가설</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">본 연구의 가설은 다음과 같이 설정하고자 한다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: -16pt; MARGIN-BOTTOM: 0pt; MARGIN-LEFT: 26pt" dir=ltr></P>
<OL style="MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt">
<LI style="BACKGROUND-COLOR: transparent; LIST-STYLE-TYPE: decimal; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none" dir=ltr>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">구속 스트레스 후 자발적 운동은 스트레스 호르몬 농도를 감소 시킬 것이다.</SPAN></P></LI>
<LI style="BACKGROUND-COLOR: transparent; LIST-STYLE-TYPE: decimal; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none" dir=ltr>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">구속 스트레스 후 자발적 운동은 노화 촉진과 관련된 단백질 발현을 억제 시킬 것이다.</SPAN></P></LI></OL>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P><BR>&nbsp;</P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: -35pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 19px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: bold; TEXT-DECORATION: none">II. 이론적 배경</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">1. 스킨스쿠버 다이빙</SPAN></P>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Morgan (1995)은 스쿠버 다이빙은 미국에서만 해마다 10만명의 다이버 당 3-9명이 사망할 정도로 위험성이 높은 스포츠 종목이며 잠수병과 관련된 질환도 매년 증가하고 있다고 하였다. 이는 심리적 불안과 공포에 의한 부분도 매우 크게 나타나며, 스트레스 반응을 감소시키기 위하여 다양한 방법의 연구가 필요하다고 하였다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Griffiths et al. (1978)은 29명의 스쿠버 다이빙을 시작하는 학생들을 대상으로 표준화된 스쿠버 테스트를 진행하기 전과 안정시의 불안 수준을 검사하였다. 안정시 특성불안과 상태불안은 평균 이하 수준이었으나, 상태불안은 스쿠버 테스트를 하는 동안 유의하게 증가하였다고 하였다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Zarezadeh &amp; Azarbanyjani (2014)는 10명의 남성 전문 다이버를 대상으로 1 m, 10 m, 20 m, 그리고 30 m의 다이빙 전 후의 채혈을 통해 스트레스 호르몬인 혈중 코티졸 농도의 변화를 살펴보았다. 혈중 코티졸은 1 m 다이빙에 비하여 모두 증가하는 것으로 나타났다. 그러나 이러한 혈청 코티졸의 변화는 오히려 불안, 심한 운동이나 추위 스트레스에 의한 것보다 증가 된 환경적 압력과 함께 침수의 생리적 효과의 결과로 판단된다고 제시하고 있다. </SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Sureda et al.(2014)은 스쿠버 다이빙이 호중구(neutrophil)의 산화적 손상(oxidative damage)과 산화질소(nitric oxide: NO) 생합성에 미치는 영향을 알아보기 위하여 9명의 다이버를 대상으로 50 m의 깊이에서 35분간 잠수하여 혈액성분을 분석하였다. 결과를 볼 때, 염증성 사이토카인이 스쿠버 다이빙 이후에 유의하게 증가하였으며, 과산화지질의 유의한 차이는 나타나지 않았다. 이러한 결과는 고농도 산소, 고압, 급성 운동 등이 결합된 상태의 스쿠버 다이빙이 nitrotyrosine 수준 증가와 nitrosative 손상 및 호중구의 염증 반응을 유도한다고 제시하고 있다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Perovic et al. (2014)은 스쿠버 다이빙 시 수중에서의 신체활동뿐만 아니라 수중에서 전개되는 다양한 환경적 요인에 의해 활성산소 생산이 증가하게 되어 산화적 손상(oxidative damage)이 매우 증가한다고 하였다. 또한, 다이버들은 스쿠버 다이빙을 하는 동안 다양한 요인에 노출됨에 의하여 혈액 동태학(haemodynamics)과 심혈관계 기능에 영향을 받게 된다고 하였다. 수중에서의 신체활동은 다이버의 심혈관계 스트레스의 주요 원인 중 한 &nbsp;가지이며, 이는 고농도의 산소, 과분압, 추위 노출, 장시간 잠수(immersion), 혈관 내의 미세기포( silent bubbles), 심리적 스트레스 등에 의해 심혈관계 기능에 영향을 주게 된다고 보고하고 있다. 또한, 제시된 요인에 단 한번 노출될 경우에도 심혈관계 불균형에 치명적인 영향을 줄 수 있다고 하였다.</SPAN></P>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none"><IMG style="BORDER-BOTTOM: medium none; BORDER-LEFT: medium none; BORDER-TOP: medium none; BORDER-RIGHT: medium none; -webkit-transform: rotate(0.00rad); transform: rotate(0.00rad)" src="https://img1.daumcdn.net/relay/cafe/original/?fname=https%3A%2F%2Flh5.googleusercontent.com%2FETfeUApAWOQXIbAYiEHftKk_uO4cKjIIIkXlZaoUchvQvgPCh9GhK8kMWBa7eftc0w28vQUeDPdHN_zC-987ZsyRD9c0-NmpthX708-HliuL-i5iqZhWLga8kfWdzNFf4w" width=556 height=350></SPAN></P>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: #ffffff; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: 'Times New Roman'; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 16px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Fig. 1. Schematic presentation of the potential connection between the increase in ROS production during dives and an increase of expression&#8206; or activation of sirtuins, along with many age-related diseases linked to the activity of sirtuins (Perovic et al., 2014).</SPAN></P>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Lambrechts et al. (2013)은 한번의 스쿠버 다이빙이 피부 의존형과 비의존형 반응성(endothelium-dependent, -independent reactivity)의 미세혈관(microvascular) 변화를 유발한다고 하였다. 이에 대한 다이빙 후의 변화를 알아보기 위하여 400 kPa에서 30분, 해수면에서 41분간 머리를 담근 채 오리발 차기(중등도 강도의 운동), 170 kPa에서 100% 산소로 41분간의 다이빙(과분압 유발)을 통한 과산소 유발 등의 환경을 통해 도플러 검사를 하였다. 결과를 볼 때 고강도로 훈련된 전문 다이버들은 한번의 다이빙으로 인해 피부 의존형 미세혈관의 변화를 보였다고 하였다. 이는 미세혈관 기능에 대한 감압의 영향이 다이빙에 순응하게 되면 바뀔 수 있다는 점을 시사한다고 하였다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Madden et al. (2014)은 스쿠버 다이빙 전 고강도의 자전거 운동이 잠수병에 미치는 영향과 극미립자(microparticle) 생성, 그리고 호중구의 활성을 알아보기 위한 연구를 하였다. 10명의 남성 다이버를 대상으로 2회의 다이빙을 행할 때, 윙게이트 사이클 프로토콜을 반복하거나 하지 않는 경우를 적용하였다. 고강도 사이클 운동을 한 결과, 다이빙 이후의 극미립자 생성이 억제되었으며, 호중구의 상승이 대조구에 비하여 억제되는 것으로 나타났다고 하였다. 이러한 결과는 스쿠버 다이빙 전에 반복적인 고강도 윙게이트 프로토콜의 적용은 극미립자 생성을 억제하므로 잠수병을 예방할 수 있을 것이라 제시하고 있다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Sureda et al. (2012)은 스쿠버 다이빙은 혈관 내 항산화 시스템을 활성화시킨다고 하였다. 이를 검토하기 위하여 9명의 남성 다이버를 수심 50 m에 35분간 잠수시켰을 때 안정시, 다이빙 직후, 다이빙 3시간 후에 채혈하여 혈액 변인을 분석하였다. 결과를 볼 때, 고수심 잠수는 과분압과 관련된 폐산소량의 증가로 인해 유발되는 산화스트레스에 대응하기 위하여 혈중 catalase, superoxide dismutase 등의 항산화시스템이 증가하였다. 다이빙은 혈관 상피세포의 상장요소를 증가시켜 다이빙에 의한 산화적 손상에 대한 저항성을 개선하는 효과를 준다고 제시하고 있다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Sureda et al. (2009)은 스쿠버 다이빙이 적혈구 및 혈중 항산화 시스템을 증가시키고 산화스트레스가 없는 상태에서 산화질소를 생산하지 않는다고 하였다. 이를 증명하기 위하여 7명의 전문 다이버를 대상으로 25분간 40 m에서 다이빙을 수행하였으며, 다이빙 전과 다이빙 직후, 3시간 후의 채혈을 진행하여 분석하였다. 결과를 볼 때, 혈중 적혈구 순환량, 헤로글로빈, 빌리루빈 등은 차이가 없었으며, 적혈구 크기가 다이빙 3시간 이후에 감소함으로서 헤마토크리트 수준이 감소하였다고 하였다. 혈중 superoxide dismutase 수준은 다이빙 이후에 유의하게 증가하였으며, 3시간 휴식에 의해 안정상태로 회귀하였다고 하였다. 아질산염(nitrite)의 수준은 혈액과 적혈구 모두 휴식기에 유의한 증가를 나타내었다. 결과적으로 스쿠버 다이빙은 혈액과 적혈구의 세포손상과 산화질소의 증가 없이 항산화 반응을 증가시킨다고 하였다. 이는 혈관 확장과 관련이 있다고 하였다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Ferrer et al. (2007)은 스쿠버 다이빙이 림프구(lymphocyte)와 호중구(neutrophil)의 항산화 방어능력이 증가한다는 것을 증명하기 위하여 7명의 남성 다이버에게 수심 40 m에서 25분간 잠수시켜 혈중 변인을 분석하였다. 또한 고압챔버에서 고압상태를 유지한 후 안정시의 변인을 분석비교 하였다. 고압챔버 상태 직후 및 다이빙 직후에 임파구의 과산화수소(H2O2) 생성이 증가하였으며, 다이빙 직후 glutathione peroxidase (GPx)와 catalase의 활성이 증가하였다. 그러나 고압챔버 상태에서는 GPx 활성만이 증가하였다. 이러한 결과를 통하여 스쿠버 다이빙에 관련된 고농도 산소는 산화적 손상을 방지하기 위해 림프구의 H2O2 &nbsp;생산과 스트레스에 의한 산화 증가를 &nbsp;항산화 효소 적응의 상태로 유도함으로서 항산화 방어체계를 개선한다고 제시하고 있다.</SPAN></P>
<P><BR><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">2. 구속스트레스</SPAN></P>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Pang et al.(2014)은 만성적인 고립스트레스가 피부의 멜라닌 생합성에 미치는 영향을 연구하기 위하여 장기간 구속스트레스 또는 간헐적인 가벼운 스트레스를 마우스에게 부하하였다. 두 가지 스트레스 모두 시상하부-뇌하수체-부신피질 축 (hypothalamic-pituitary-adrenocortical (HPA) axis)의 mRNA 발현 감소를 유발하였으며, 멜라닌 생합성을 저해하였다고 보고하였다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><A style="TEXT-DECORATION: none" href="http://www.ncbi.nlm.nih.gov/pubmed?term=Wis%C5%82owska-Stanek%20A%5BAuthor%5D&amp;cauthor=true&amp;cauthor_uid=25344674"><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Wisłowska-Stanek</SPAN></A><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none"> et al. (2013)은 만성 구속 스트레스에 노출된 쥐의 경우, 해마와 대뇌 피질 GABAergic 신경 전달의 기능이 약화되고, 이 효과는 시상 하부 - 뇌하수체 - 부신 축의 기능과 우울 증상에 중요한 영향을 미칠 수 있다고 하였다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Jin et al. (2014)은 급성 구속스트레스는 Gsk-3β의 활성화를 유도하여 장기간 강화(long-term potentiation)되는 과정을 손상시킨다고 하였다. 이를 증명하기 위하여 마우스를 1시간 동안 구속시킨 후에 해마회(hippocampal)를 절편하여 분석하였다. 결과를 볼 때, Ser473의 Akt 인산화가 증가되었으며, Ser9의 Gsk-3β의 인산화가 발견되었으나, Tyr216에서는 발견되지 않았다고 하였다. 이러한 결과는 장기간 강화의 손상이 Akt 활성화의 억제에 의한 Gsk-3β 활성화 개선에 의해 유발된다고 하였다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Maghsoudi et al. (2014)은 임신한 쥐에게 7일간 하루 1시간 또는 3시간 동안 만성적인 고립 스트레스를 부하하였을 때 나타난 결과를 살펴보았다. 결과를 볼 때, ACTH (adrenocorticotropic hormone) 및 corticosterone 농도가 증가하였으며, BDNF (brain-derived neutrophic factor)와 신경생장인자 수용체의 일종인 TrkB 단백질 발현이 3시간의 고립 스트레스를 부하한 경우에 유의하게 감소하였다고 하였다. 이러한 결과를 통하여 임신 중 스트레스 상황은 우울증과 유사한 행동 반응을 증가시킬 수 있다고 제시하고 있다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Fair et al. (2014)은 포획되었다가 방류되는 병코 돌고래(큰돌고래, Bottlenose dolphins)의 스트레스 반응에 대하여 연구하였다. 포획되었을 때 샘플 채취의 평균 시간은 22분이었으며, 방류되기까지 평균 1시간 37분이 경과되었을 때의 2회 채혈을 통하여 스트레스 호르몬 변화를 분석하였다. 에피네프린과 도파민 농도는 최초 포획되었을 때에는 차이가 없었으나, 시간이 경과할 수록 점차 증가하였으며, 코티졸와 ACTH 농도는 최초 포획시부터 방류시점까지 지속적으로 증가한 상태로 유지되었다. 스트레스 반응은 어린 돌고래가 어른 돌고래에 비하여 높게 나타났다. 이러한 결과들을 통하여 야생 돌고래 포획과 구속의 급성 스트레스에 대한 전형적인 포유류의 반응을 보였다고 보고하고 있다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Jameel et al. (2014)은 Wistar 쥐를 이용하여 강제수영운동과 구속 상황에서의 혈중 코티졸 농도의 변화를 살펴보았다. 결과를 볼 때, 혈중 코티졸 농도는 강제수영운동과 마찬가지로 구속 상태에서도 유의하게 증가하였다. 구속 상태 이후에 강제수영운동을 추가로 부하하였을 경우에는 유의하지는 않았으나 코티졸 농도의 지속적인 상승이 관찰되었다. 이러한 결과는 구속 상태가 강제수영운동에 비하여 더욱 많은 스트레스를 주는 방법이라고 제시하고 있다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Shirasaki et al. (2013)은 키노몰구스 원숭이(Cynomolgus monkey)를 의자에 묶어 놓은 상태의 신체활동을 제한하였을 때 나타나는 글루코스 내성을 분석하였다. 결과를 볼 때, 구속 스트레스는 혈중 글루코스 농도를 증가시키며 코티졸 농도를 증가시킨다고 하였다. </SPAN></P>
<P><BR><BR>&nbsp;</P>
<P style="TEXT-ALIGN: justify; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">3. 운동과 스트레스</SPAN></P>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Han et al. (2014)은 난소를 적출한 쥐에게 21일간 반복적인 고립스트레스 후 장기간 트레드밀 달리기 운동훈련을 부하하였다. 그 후 강제수영운동을 시킨 결과, &nbsp;트레드밀 운동훈련에 의해 호르몬 농도가 개선되었음을 보고하였다. 이는 폐경 이후에 나타날 수 있는 우울증이 장기간 운동에 의해 개선될 수 있다는 점을 제시하고 있다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Diverio et al. (2014)의 눈사태 구조견(avalanche dogs)을 활용한 연구에서는, 인공적으로 눈사태를 만든 지역에서 사람이 묻힌 상태를 만든 후, 탐지 활동을 하였을 때 조난된 사람을 발견한 시점에서 스트레스와 관련된 유전자 활성이 가장 높게 나타났다고 하였다. 이는 일회성 운동을 하였을 때와 동일하며, 사람 발견 후 2시간이 경과되었을 때 다시 감소하는 결과를 보였다고 하였다. </SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Uribe et al. (2014)은 Wistar 쥐에게 최대 14일간의 자발적 운동, 18%의 식이제한 또는 반복적인 구속스트레스를 부하한 후 에너지 항상성에 영향을 미치는 &nbsp;hypothalamic-pituitary thyroid (HPT) 축(axis)의 활성을 조사하였다. 자발적 운동군에서는 HPT 축의 활성이 억제되는 것이 방지되었으나, 반복적인 구속스트레스군에서는 활성이 억제되었다. 이는 자발적인 운동이 반복적인 스트레스 혹은 식이제한에 의한 대사경로의 변화를 억제할 수 있다고 제시하고 있다. </SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Castella-Ortega et al. (2014)는 만성적인 스트레스가 유발되었을 때, 자발적인 운동이 에피소드 같은 기억력과 성숙한 해마의 신경생합성에 미치는 영향을 알아보고자 하였다. 만성적인 스트레스 작용 방법은 간헐적인 구속스트레스로 하였다. 결과적으로 스트레스 이후에 자발적인 운동을 병행하였을 경우, 스트레스가 경감되었으며, 기억력과 성숙된 해마의 신경생합성 역시 개선되었다고 하였다. 이러한 결과는 지속적인 스트레스를 받고 있는 경우에도 자발적인 운동에 참여할 경우 운동에 의해 해마의 신경생합성이 개선된다고 제시하고 있다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Luo et al. (2014)은 구속스트레스가 장관 기능과 항균 작용을 억제시킨다는 것에 착안하여 중등도 수영운동훈련을 병행하였을 때 나타나는 변화를 살펴보았다. 구속스트레스 30분 전에 하루 30분간의 수영운동훈련은 장내세균총의 이동을 촉진시켰으며, 장내 투과성을 유지시키는 것으로 나타났다. 이러한 결과는 중등도 강도의 운동훈련이 만성적인 구속스트레스로 인한 장관 기능 손상이 억제되었으며, 이는 소장에서의 항균작용이 유발되었기 때문이라고 제시하고 있다.</SPAN></P>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none"><IMG style="BORDER-BOTTOM: medium none; BORDER-LEFT: medium none; BORDER-TOP: medium none; BORDER-RIGHT: medium none; -webkit-transform: rotate(0.00rad); transform: rotate(0.00rad)" src="https://img1.daumcdn.net/relay/cafe/original/?fname=https%3A%2F%2Flh6.googleusercontent.com%2F2jeYEwtJBmCaS2waigT-g6USCAyBkrEj3LJsGcFHC2sdOdbhpUHpEoZQ1zswufvVU3a_siqnc_752pw_WnIhkIf1y4Zmvn6EIF7Hpsqkm_NdnXexDPmRu4BSQAT9UFprNw" width=448 height=272></SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: #ffffff; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: 'Times New Roman'; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 16px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Fig. 2. Latency of entrance to the dark compartment of passive avoidance apparatus during memory retention test 21 days after receiving foot shock. Data represent mean ± SEM (n=10). Co/Sh groups vs. Ex, St and St&amp;Ex groups, in all of them p&gt;0.05 and nonsignificant; Ex group vs. St group, ‡ P&lt;0.05; St group vs. St &amp; Ex groups, P&gt;0.05 and nonsignificant; Kruskal -Wallis test, Mann-Whitney U test. Co: Control group; Sh: Sham group; Ex: Exercise group; St: Stress group; St&amp;Ex: Synchronized exercise with stress group. ‡ P&lt;0.05 with respect to the Exercise group (Radahmadi et al., 2013).</SPAN></P>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Radahmadi et al. (2013)은 학습능력과 기억의 손상이 운동에 의해 개선될 수 있는지를 알아보가 위하여 만성적인 스트레스가 유발된 쥐에게 트레드밀 달리기를 부하하여 조사하였다. 만성적인 구속스트레스는 하루 1시간씩 21일간 지속하였다. 결과적으로 스트레스가 없는 상태에서 운동만을 하였을 때는 단기, 중기의 기억이 개선되었으나, 구속스트레스 자극과 트레드밀 달리기 운동을 병행하였을 경우에는 아무런 개선효과를 발견하지 못하였다고 하였다. 오히려 구속스트레스만을 유발시킨 쥐와 마찬가지로 기억력의 손상이 발견되기도 하였다. 이러한 결과는 강제로 달리기 운동을 부하하였기 때문이라고 제시하고 있다.</SPAN></P>
<P><BR><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">4. 자발적 운동과 스트레스</SPAN></P>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Uysal et al. (2014)은 자발적 운동과 강제운동을 부하하였을 때 인지기능과 신경생합성에 영향을 미치는 VEGF 및 BDNF 수준을 조사하였다. 이를 위하여 자발적 운동 쥐에게는 6주간 자율적으로 운동을 할 수 있도록 하였으며, 강제운동 쥐에게는 6주간 주당 5일의 빈도로 분당 8 m의 속도로 30분간 운동을 부하하였다. 해마의 VGEF 및 BDNF 수준은 두 운동군 모두 증가하였으나, 자발적 운동군이 더욱 증가하였다. 또한 수컷 쥐의 경우, 불안 수준은 두 가지 운동군 모두 차이가 없었으나, 암컷 쥐의 경우 강제운동을 부하하였을 경우에 불안수준이 높아졌으며, 전두엽 피질 (prefrontal cortex) VEGF 및 BDNF 수준이 감소하였다고 보고하였다. </SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Conner et al. (2014)은 강제 운동에 대한 대안으로서 쇼크가 없는 상태에서 쥐의 지구성 운동능력을 평가하고자 하였다. 이를 위하여 상대적으로 온순한 C57BL/6 마우스를 이용하여 전기자극이 없는 상태에서의 트레드밀 운동을 부하하였다. 마우스는 각각의 쥐에 맞는 보행 속도로서 트레드밀을 자발적으로 운동하게 하였다. 이러한 방법의 적용을 통하여 실험상황에서의 스트레스에 의해 오염될 수 있는 실험결과를 보다 정교화 시킬 수 있다고 제시하였다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Ogbonmwan et al. (2014)은 코카인에 중독되었을 때 자발적 운동의 참여가 금욕 스트레스에 의해 유발된 코카인 추구를 억제할 수 있는 지에 대하여 연구하였다. 운동 쥐는 하루 4-6 km를 달렸으며, 코카인 흡입 종료 후 운동은 코카인을 요구하는 정도는 감소하였으나, 전기충격에 의한 달리기를 한 경우에는 그렇지 않았다고 하였다. 이러한 결과는 자발적인 운동 프로그램이 &nbsp;약물 등의 중단에 의해 나타나는 금단현상을 억제하고 금욕을 유지하는데 도움을 줄 수 있을 것이라 제시하고 있다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Lertsinthai et al. (2014)은 난소를 적출한 쥐의 자발적인 휠 러닝(wheel running)이 스트레스에 의해 유발된 골감소를 완화하는지를 검토하였다. 이를 위하여 만성적인 혐오 자극(chronic aversive stimuli (CAS)에 의한 스트레스를 4주간 부하한 후 자발적 휠 러닝을 부하하였다. 이때 골미네랄 함량(bone mineral content: BMC)와 골밀도(bone mineral density)를 분석하였다. 결과를 볼 때, CAS는 골소주의 골량을 감소시켰으며, 골소주의 분리를 증가시켰다. 그러나 자발적 휠 러닝을 부하하였을 경우에는 난소 적출 시기 전의 골량으로 회복되었다. 난소 적출 쥐에게 CAS에 의한 골량의 감소는 파골세포에 의한 골흡수(bone resorption)이 증가되었기 때문으로 판단된다고 하였다. 또한, 자발적 휠 러닝은 충격이 없는 운동으로서 BMD와 BMC를 증가시키는 것으로 나타났다고 하였다. </SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><A style="TEXT-DECORATION: none" href="http://www.ncbi.nlm.nih.gov/pubmed?term=Andreev-Andrievskiy%20AA%5BAuthor%5D&amp;cauthor=true&amp;cauthor_uid=25358851"><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Andreev-Andrievskiy</SPAN></A><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none"> et al. (2014)은 트레드밀 달리기를 하였을 때 실험 쥐의 심박수가 단계적으로 증가하지 않고 매우 급격한 상승을 한다는데에 착안하여 트레드밀 달리기와 자발적 휠 러닝 상태를 비교하였다. 상대적으로 온순한 C57BL/6 마우스를 이용하여 스트레스 상황을 유도한 뒤 트레드밀 달리기 혹은 자발적 휠 러닝을 부하하였다. 결과를 볼 때 사육 케이지에 설치된 자발적 휠에서 달리기를 하였을 때 심박수는 실험 쥐의 달리기 속도가 증가와 동일한 선형으로 증가하였다. 이러한 결과는 트레드밀 달리기가 일종의 스트레스와 관련된 교감 신경 활성화에 의해 주로 지배된다는 점을 시사하고 있다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Li et al. (2014)은 비자발적인 운동이 자발적인 운동에 비하여 더욱 많은 양의 신체적, 정신적 스트레스를 받는다고 하였다. 이를 검증하기 위하여 휠과 트레드밀을 활용하여 자발적인 운동과 강제운동을 부하한 후 해마의 뇌파, 심전도 등을 무선 장치를 이용하여 분석하였다. 결과를 볼 때, 트레드밀 운동을 부하하였을 경우에는 자발적인 운동에 비하여 뇌파 중 세타파(theta wave)와 &nbsp;심박수가 더 많은 변화를 보였다. 이러한 결과를 통하여 자발적인 운동에 비하여 강제운동은 주행시 추가적인 감정상태 혹은 감각상호작용의 효과를 줄 수 있다고 하였다. </SPAN></P>
<P><BR><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">5. 운동과 세포사멸</SPAN></P>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Chang et al. (2014)은 Heat shock protein 72 (HSP 72)이 운동에 의해 발현이 증가하게 되는 대표적인 스트레스 단백질로 활용되는 마커로서 척수손상(spinal cord injury: SCI) &nbsp;모델 쥐를 운동시켰을 때 높은 수준의 HSP 72가 신경에서 발견되었다고 하였다. 이러한 결과를 통하여 운동을 부하하지 않은 실험 쥐에 비하여 낮은 수준의 기능 저하의 결핍, &nbsp;적은 척수의 타박상(contusions), 세포사멸의 억제가 나타났다고 보고하였다. </SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><A style="TEXT-DECORATION: none" href="http://www.ncbi.nlm.nih.gov/pubmed?term=Mokhtari-Zaer%20A%5BAuthor%5D&amp;cauthor=true&amp;cauthor_uid=24906198"><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Mokhtari-Zaer </SPAN></A><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">et al. (2014)은 자발적인 운동 또는 트레드밀 운동이 몰핀에 의존하는 쥐의 세포사와 관련된 단백질 발현에 변화를 가져온다고 하였다. 이를 증명하기 위하여 가벼운 트레드밀 달리기 또는 자발적 휠 러닝을 10일간 부하하였다. 결과를 볼 때, 만성적인 몰핀 투여는 Bcl-2의 발현을 지속적으로 억제하였으나 두 가지 운동부하 상태 모두 반대의 결과가 나타났다. 결과적으로 약물 중독 등에 의한 세포사멸의 증대는 지속적인 운동에 의해 개선될 수 있으며, 약물 중독자들의 치료 전략으로 제시될 수 있다고 하였다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Ding et al. (2014)은 고강도 운동이 과도한 세포사멸을 유도한다는 것을 증명하기 위하여 중추신경계에 미치는 영향을 살펴보았다. 이를 위하여 1회의 수영을 탈진시까지 부하하여 24시간이 경과되었을 때 나타난 결과를 살펴보았다. 결과를 볼 때, caspase-3, Bax, caspase-12 등의 분열이 증가하였으며, Bcl-2는 감소한 것으로 나타났다. </SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><A style="TEXT-DECORATION: none" href="http://www.ncbi.nlm.nih.gov/pubmed?term=Mour%C3%A3o%20FA%5BAuthor%5D&amp;cauthor=true&amp;cauthor_uid=25063943"><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Mour&atilde;o</SPAN></A><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none"> et al. (2014)은 운동이 허혈손상(ischemic injury)에 대한 신경보호 역할을 하게 되지만, 운동이 뇌의 허혈에 미치는 영향에 대해서 연구가 부족하다는데에 착안하여 이를 검증하고자 하였다. 이를 위하여 8주간 수영운동을 부하하여 해마 조직을 연구하였다. 결과를 볼 때, 세포 사멸율, 세포사 마커의 발현이 운동에 의해 현저하게 감소하였다. 이는 글루탐산성 흥분독성(glutamatergic excitotoxicity) 및 세포사멸의 감소에 의한 것이라 제시하고 있다. 또한 수영훈련에 의해 세포사멸 마커인 caspase-8, 9, 3 등이 감소하였다고 보고하였다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Kim et al. (2014)은 당뇨 쥐에게서 태어난 어린 쥐가 트레드밀 운동을 하였을 때, 해마의 세포 분화 개선, 세포사멸의 억제 등으로 인하여 단기 기억 손상을 완화시킨다고 하였다. 이를 증명하기 위하여 어미 쥐에게 streptozotocin에 의한 당뇨를 유발한 후 교미시켜 태어난 새끼 쥐가 4주령이 되었을 때 2주간 트레드밀 운동을 하루 30분씩 부하하여 해마 조직에서 나타난 변화를 살펴보았다. 결과를 보면, 트레드밀 운동에 의해 세포 분화가 개선되었으며, 해마의 세포사멸이 억제된 것으로 나타났다. 이는 선천성 당뇨를 가지고 있는 어린이들에게 트레드밀 운동이 신경발달 문제를 개선시킬 수도 있다는 점을 시사한다고 하였다.</SPAN></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none"><IMG style="BORDER-BOTTOM: medium none; BORDER-LEFT: medium none; BORDER-TOP: medium none; BORDER-RIGHT: medium none; -webkit-transform: rotate(0.00rad); transform: rotate(0.00rad)" src="https://img1.daumcdn.net/relay/cafe/original/?fname=https%3A%2F%2Flh3.googleusercontent.com%2Fu3BQFOQuz0oIkifPPydIvLHHeRnT72SRFZBoqwiAbhOFtjo1jzsrB5jVsNmrVYbWcD_Qs7m2Swg6Owgmt58rQ_BGBvCOm-4VCOfD4taVnKlmdAQKYukXkj0hy8jnF9XLTg" width=370 height=330></SPAN></P>
<P style="LINE-HEIGHT: 1; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: #ffffff; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: 'Times New Roman'; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 16px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Fig. 3. Apoptotic signaling pathways. Schematic overview of three major caspases-dependent apoptotic signaling pathways including (a) cytokines/Fas-mediated caspase-8 pathways, (b) mitochondrial-mediated caspase-9 pathways, and (c) endoplasmic reticulum (ER)/Ca2+-mediated caspase-12 pathways. (Kwak, 2013).</SPAN></P>
<P><BR>&nbsp;</P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none"><IMG style="BORDER-BOTTOM: medium none; BORDER-LEFT: medium none; BORDER-TOP: medium none; BORDER-RIGHT: medium none; -webkit-transform: rotate(0.00rad); transform: rotate(0.00rad)" src="https://img1.daumcdn.net/relay/cafe/original/?fname=https%3A%2F%2Flh6.googleusercontent.com%2FUzYnIek9lRRtR7UnPNoRJqs28PJs2Ggj1yZm4oMoPKZNVIZbdxOwiBKcCmYi2q4g3T-F0iRCtCLpA7WNAql4xWgi_l0IXVP7GFXaSmBhnzROvChcP9zTEFiMmRM8OEjnrQ" width=349 height=298></SPAN></P>
<P style="LINE-HEIGHT: 1; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: #ffffff; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: 'Times New Roman'; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 16px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Fig. 4. Exercise protection model against apoptosis in the heart. An integrative model of exercise protection against apoptosis in the heart whereby exercise can potentially upregulate stress proteins such as MnSOD, ERK, IGF-1/Akt, HSPs, and NF-kB (Kwak, 2013).</SPAN></P>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Fig 3과 Fig 4 (Kwak, 2013)에서 제시하고 있는 바와 같이, 세포사멸을 유발하는 신호전달 체계는 매우 다양하게 나타난다. 그러나, 운동을 부하하였을 때 세포사멸을 억제할 수 있는 것으로 나타난다.</SPAN></P>
<P><BR><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 19px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: bold; TEXT-DECORATION: none">III. 연구방법</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">1. 실험동물 및 사육방법</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">본 연구에서는 생후 5주령의 수컷 ICR 마우스 21마리를 사용할 것이며, 마우스는 케이지당 4마리 또는 3마리로 분류하고, 사육실 내부 환경은 자동 냉난방기를 이용하여ㅠ23-25℃, 상대습도는 60% 내외를 유지시키고 08:00~20:00까지 12시간 간격으로 명&#8228;암주기를 조절할 것이다. 집단구분은 대조군(Control; CON), 구속스트레스군(Restraint Stress; RES), 구속스트레스 + 자발적운동군(Restraint Stress + Voluntary Wheel Running; RESVE)으로 구분 할 것이며, 집단구분은 &lt;Table 1&gt;과 같다. 본 연구는 경북대학교 실험동물윤리위원회의 승인 후 진행할 것이다.</SPAN></P>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Table 1. Division of experimental rats</SPAN></P>
<DIV style="MARGIN-LEFT: 0pt" dir=ltr>
<TABLE style="BORDER-BOTTOM: medium none; BORDER-LEFT: medium none; BORDER-COLLAPSE: collapse; BORDER-TOP: medium none; BORDER-RIGHT: medium none">
<COLGROUP>
<COL width=185>
<COL width=185>
<COL width=209></COLGROUP>
<TBODY>
<TR style="HEIGHT: 0px">
<TD style="BORDER-BOTTOM: #000000 3px solid; BORDER-LEFT: #000000 0px solid; PADDING-BOTTOM: 1px; PADDING-LEFT: 1px; PADDING-RIGHT: 1px; VERTICAL-ALIGN: middle; BORDER-TOP: #000000 3px solid; BORDER-RIGHT: #000000 0px solid; PADDING-TOP: 1px">
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Control</SPAN></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">(CON)</SPAN></P></TD>
<TD style="BORDER-BOTTOM: #000000 3px solid; BORDER-LEFT: #000000 0px solid; PADDING-BOTTOM: 1px; PADDING-LEFT: 1px; PADDING-RIGHT: 1px; VERTICAL-ALIGN: middle; BORDER-TOP: #000000 3px solid; BORDER-RIGHT: #000000 0px solid; PADDING-TOP: 1px">
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Restraint Stress</SPAN></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">(RES)</SPAN></P></TD>
<TD style="BORDER-BOTTOM: #000000 3px solid; BORDER-LEFT: #000000 0px solid; PADDING-BOTTOM: 1px; PADDING-LEFT: 1px; PADDING-RIGHT: 1px; VERTICAL-ALIGN: middle; BORDER-TOP: #000000 3px solid; BORDER-RIGHT: #000000 0px solid; PADDING-TOP: 1px">
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Restraint Stress +</SPAN></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Voluntary Wheel Running</SPAN></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">(RESVE)</SPAN></P></TD></TR>
<TR style="HEIGHT: 0px">
<TD style="BORDER-BOTTOM: #000000 3px solid; BORDER-LEFT: #000000 0px solid; PADDING-BOTTOM: 1px; PADDING-LEFT: 1px; PADDING-RIGHT: 1px; VERTICAL-ALIGN: middle; BORDER-TOP: #000000 3px solid; BORDER-RIGHT: #000000 0px solid; PADDING-TOP: 1px">
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: #ffffff; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">n=7</SPAN></P></TD>
<TD style="BORDER-BOTTOM: #000000 3px solid; BORDER-LEFT: #000000 0px solid; PADDING-BOTTOM: 1px; PADDING-LEFT: 1px; PADDING-RIGHT: 1px; VERTICAL-ALIGN: middle; BORDER-TOP: #000000 3px solid; BORDER-RIGHT: #000000 0px solid; PADDING-TOP: 1px">
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: #ffffff; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">n=7</SPAN></P></TD>
<TD style="BORDER-BOTTOM: #000000 3px solid; BORDER-LEFT: #000000 0px solid; PADDING-BOTTOM: 1px; PADDING-LEFT: 1px; PADDING-RIGHT: 1px; VERTICAL-ALIGN: middle; BORDER-TOP: #000000 3px solid; BORDER-RIGHT: #000000 0px solid; PADDING-TOP: 1px">
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 1.15; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: #ffffff; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">n=7</SPAN></P></TD></TR></TBODY></TABLE></DIV>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">2. 식이조성 및 섭취방법</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">사료는 효창사이언스(formura-m07, 제일사료, 한국)에서 펠렛형태의 사료를 공급받아 사용 할 것이며, 식이와 물은 자유섭취 할 것이다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">3. 구속스트레스 방법</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">구속스트레스는 강제 수영운동에 비하여 더 큰 스트레스를 부하하는 방법이다(Jameel et al., 2014). 아래 Fig. 5와 같이, 마우스를 움직이지 못하게 한 상태에서 30 m/d (Fan et al. 2013)로서 4주간 진행하고자 한다.</SPAN></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none"><IMG style="BORDER-BOTTOM: medium none; BORDER-LEFT: medium none; BORDER-TOP: medium none; BORDER-RIGHT: medium none; -webkit-transform: rotate(0.00rad); transform: rotate(0.00rad)" src="https://img1.daumcdn.net/relay/cafe/original/?fname=https%3A%2F%2Flh4.googleusercontent.com%2FQe2JPPc_QLrPkzAYi99mD7FZOEsdmLWB734pmVcyVl_ncz6dwqSbCppvzUrse_CXEHflhTQF-dchqnHZJqHl71BCFdWp5_uMSNH8B2ksUiTZlGe3Myj8j_R45L-aRl6URg" width=431 height=218></SPAN></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Fig. 5. Restraint stress model</SPAN></P>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">4. 자발적 운동방법</SPAN></P>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">예비사육 후 6주령이 되었을때 부터 쳇바퀴를 24시간 동안 자발적 운동을 4주간 매일 시행 할 것이다. 실험 기간 중 매일 오전 9시에 자발적 운동량 체크 후 reset을 할 것이다. 운동장치는 Fig. 6과 같다. 운동장치의 지름은 370 mm으로서 한 바퀴 회전하였을 때 거리는 1162.4 mm 즉, 1.1624 m 이다. 사육장소에서 운동장치의 입구는 통행이 가능하지만, 외부로의 통행은 불가능하게 구성되어 있어 운동을 하고 싶을 경우에는 항상 운동장치에 진입하여 운동을 할 수 있게 되어 있다. 또한, 운동장치는 한 바퀴 회전할때 마다 센서에 의해 회전 수가 자동으로 기록되도록 설계하였다.</SPAN></P>
<P><BR>&nbsp;</P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none"><IMG style="BORDER-BOTTOM: medium none; BORDER-LEFT: medium none; BORDER-TOP: medium none; BORDER-RIGHT: medium none; -webkit-transform: rotate(0.00rad); transform: rotate(0.00rad)" src="https://img1.daumcdn.net/relay/cafe/original/?fname=https%3A%2F%2Flh3.googleusercontent.com%2F1y_aZ7tVP0DGwXGk8z_CJsdtt6m73-qY8Uakz7ECfYp4iveWtDCy1KuJSeSwoiJFpYbklpkyd_iSH0XaSRpicz_Gs1kgWqZp8lt2SVt1k0UTuyvjpW1RDvBrORa6mavS7w" width=354 height=218></SPAN></P>
<P style="TEXT-ALIGN: center; LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">Fig. 6. Voluntary exercise training apparatus</SPAN></P>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">5. 시료채취</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">12시간 절식 후 lsoflurane (Forane, Aesioa Queen Borough Ltd, UK)의 흡입 마취 (Srgivet , Isotec , USA) 상태에서 개복하여, 복대동맥에서 채혈할 것이다. 채취된 혈액은 3,000 rpm으로 10분간 원심분리 (1580MER, Gyrozen, Korea) 후 분석 시까지 냉동고 (NF-400SF, HFC, Japan) 에 보관할 것이다. 그 후 간과 근육 등 분석에 필요한 장기를 적출하여 &nbsp;액체 질소를 활용하여 &nbsp;활성을 정지시킨 상태에서 분석시 까지 &nbsp;-80℃ 냉동고 (NF-400SF, HFC, Japan)에 분석 시 까지 보관할 것이다.</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">6. 분석항목 및 방법</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<OL style="MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt">
<LI style="BACKGROUND-COLOR: transparent; LIST-STYLE-TYPE: decimal; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none" dir=ltr>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">체중증가율</SPAN></P></LI></OL>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">체중의 측정은 동일한 시간과 환경에서 매주 2회 측정 할 것이다. 즉, 실험 기간 중 매주 월요일, 목요일 오전 9시에 규칙적으로 측정하여 동일한 환경을 유지할 수 있도록 할 것이다. &nbsp;측정된 주당 2회의 자료를 평균내어 주당 변화 그래프를 작성하여 표기할 것이다.</SPAN></P>
<P><BR>&nbsp;</P>
<OL style="MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" start=2>
<LI style="BACKGROUND-COLOR: transparent; LIST-STYLE-TYPE: decimal; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none" dir=ltr>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">운동량</SPAN></P></LI></OL>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">운동량 측정은 4주동안 매일 측정 할 것이다. 실험 기간 중 매일 오후 9시에 24시간 자발적 운동량을 체크 후 reset을 할 것이다. 측정된 운동량은 평균을 내어 주당 변화그래프를 표기할 것이다.</SPAN></P>
<P><BR>&nbsp;</P>
<OL style="MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" start=3>
<LI style="BACKGROUND-COLOR: transparent; LIST-STYLE-TYPE: decimal; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none" dir=ltr>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">지방량 측정</SPAN></P></LI></OL>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">복강내 지방과, 고환지방을 정량하기 위하여 실험쥐를 눕힌 상태에서 복부를 개복한다. &nbsp;복강내에 있는 지방과 고환에 있는 지방을 적출하여 정량화 한다. &nbsp;</SPAN></P>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">1) 혈중 스트레스 호르몬</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">혈청의 cortisol 농도는 cortisol ELISA Kit를 이용하여 분석 할 것이다. 채취한 혈액 샘플을 1000 rpm에서 10분간 원심분리 후 &nbsp;상온에서 20분 방치 한다. 방치한 샘플은 kit의 희석법에 따라 미리 희석한 실험샘플과 Standard를 각각의 Well에 100 ㎕ 분주한 후 37 ℃ shaker에서 90분동안 혼합한다. &nbsp;그 후 wash buffer를 이용하여 3번 세척하되 각각의 well에 400 ㎕를 분주한 후, plate를 거꾸로 하여 wash buffer를 제거한다. 각 well에 cortisol antibody를 100 ㎕를 분주 한 후 37 ℃ shaker에서 60분동안 혼합한다. &nbsp;그리고, wash buffer를 이용하여 3번 세척한다. cortisol enzyme-conjugate 100 ㎕ 분주한 후, 37 ℃ shaker에서 30분동안 혼합한다. 마지막으로 wash buffer 400㎕ 를 이용하여 5번 세척하되 뒤집어서 tapping 한다. 세척 후 Colour Reagent 100 ㎕를 분주 후 &nbsp;어두운 shaker에서 잘 혼합 한다. 혼합 되었던 plate의 well에 Colour Reagent C를 100 ㎕분주 하고 가볍게 혼합하여준 후 30분 이내에 ELISA reader를 이용하여 45nm에서 흡광도를 측정한다.</SPAN></P>
<P><BR>&nbsp;</P>
<OL style="MARGIN-TOP: 0pt; MARGIN-BOTTOM: 0pt" start=2>
<LI style="BACKGROUND-COLOR: #ffffff; LIST-STYLE-TYPE: decimal; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none" dir=ltr>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; MARGIN-BOTTOM: 3pt" dir=ltr><SPAN style="BACKGROUND-COLOR: #ffffff; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">단백질 발현</SPAN></P></LI></OL>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">근육 조직과 간 조직의 처리는 RIPA Buffer (RIPA Buffer, Biosesang, Korea) 1ml을 샘플 0.1g에 넣어 균질화 작업을 할 것이다. 균질화 작업이 끝난 후 샘플을 12,000 rpm 에서 10분간 원심분리 후 상층액을 분리 한 다음, 각 샘플의 단백질 정량을 하기위하여 분리한 상층액 1 ㎕와 5 x Brad-ford (Protein Assay, Bio-Rad, USA) &nbsp;1 ml을 혼합 후 각 샘플을 ELISA plate (TECAN, Austria)에 200 ㎕를 넣어서 595 nm의 파장으로 흡광도를 측정한 다음 3 x SDS-PAGE sample buffer를 섞어서 샘플을 만들어 100℃에서 10분간 끓인 후 ice에 넣어 놓는다. 단백질 분석을 위하여 10% SDS-PAGE에 protein marker (Xpert2 prestained protein marker, Gindepot, USA) 10 ㎕와 단 백질 샘플 15 ㎕를 loading한 후에 80 V에서 전기영동하여 단백질을 분리한다. 단백질 분리 후 polyvinylidene difluoride membranes (Amersham Hybond, GE Healthcare, USA) 으로 electroblotting 100 V에 의해 60분간 membrane으로 전이시킨다. 전이가 끝난 후 skim milk (Skim milk powder, MB cell, Korea) 가 포함 된 blocking buffer (TBST buffer; 50 mM Tris-Hcl; 150 mM NaCl; 0.05%, hnology Inc, sc-9117, USA) , Hsp70: 1:2000(abcam, ab94368, UK), Bcl-2: 1:200(Santa cruz biotechnology Inc, sc-49, USA), p53:1:1000(abcam, ab26, UK), &nbsp;Caspase-3: 1:1000 (Cell Signaling Technolohy Inc, #9665, USA), 각각 방법에 따라 희석한 후 4℃에서 over-night 시킨 후 TBST(5% tween-20) 를 이용하여 10분간 5회 washing 한다. 2차 항체는 &nbsp;Hsp70: 1:5000, Bcl-2: 1:3000, Caspase-3: 1:2000(goat-anti-rabbit, Santa cruz Biotechnology, Inc, CA NO. sc-2004, USA), p53:1:5000(goat anti mouse, Santa cruz Biotechnology, Inc, CA NO. sc-2005, USA) 를 각각에 방법에 따라 희석하여 두 시간 동안 반응을 시킨 후 TBST (5% tween-20) 로 10분간 5회 washing 한다. 이에 따라 도출된 근육에 대한 밴드의 동정은 enhanced-chemilumine scene (</SPAN><SPAN style="BACKGROUND-COLOR: #ffffff; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">RPN 2106, </SPAN><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">GE Healthcare, USA) kit을 이용해 membrane을 발광 시킨 후 X-ray flim (Ortho-CP-G plus, AGFA, Belgium) 에 현상한다. </SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">현상 후, B-actin은 1:3000 (Santa cruz biotechnology Inc, sc CA NO. sc-47778, USA) 을 각각에 방법에 따라 희석하여 4℃에서 over-night 시킨 후 TBST(5% tween-20) 를 이용하여 10분간 5회 washing 한다. 2차 항체는 goat anti rabbit 1:3000 (Santa cruz biotechnology Inc, sc CA NO. sc-2005, USA) 을 각 방법에 따라 희석하여 두 시간동안 반응을 시킨 후 TBST로 10분간 5회 washing 한 후 ECL kit을 이용해 membrane을 발광 시킨 후 X-ray flim 에 현상한다. band의 density는 image j(NIH ver. 1.48, USA)를 이용하여 band/β-actin으로 계산한다.</SPAN></P>
<P><BR>&nbsp;</P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">7. 자료처리 방법</SPAN></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr></P>
<P style="LINE-HEIGHT: 2; MARGIN-TOP: 0pt; TEXT-INDENT: 10pt; MARGIN-BOTTOM: 0pt" dir=ltr><SPAN style="BACKGROUND-COLOR: transparent; FONT-VARIANT: normal; FONT-STYLE: normal; FONT-FAMILY: Arial; WHITE-SPACE: pre-wrap; COLOR: #000000; FONT-SIZE: 15px; VERTICAL-ALIGN: baseline; FONT-WEIGHT: normal; TEXT-DECORATION: none">연구결과의 데이터는 Windows용 통계프로그램 SPSS/PC+21.0을 이용한다. 모든 실험 결과는 평균과 표준오차로 나타내고, 각 그룹의 유의성 검증을 위하여 일원분산분석을 실시하며, 유의한 차이가 나타난 항목에 대해서는 least significant different 법으로 사후검정을 실시한다. 모든 차의 유의성은 p&lt;.05 이하로 설정하고자 한다.</SPAN></P>
<P><BR><BR></B></P>&nbsp;</P>
<p>&nbsp;</p>
<!-- -->
카페 게시글
검색이 허용된 게시물입니다.
살아가는 이야기들
13일 졸업논문 계획서 발표입니다.
다음검색




첫댓글 오우... 동물학대요.. 피뇨형... 형의 지적인 모습에는 정말 깜놀 한답니다. 축하드려요.. 피뇨박사님.
참 글고 이제 2014년도 다 끝나가는데.. 저 알바비 입금은 언제쯤이나... ㅎㅎㅎ 게시글도 많이 올렸고.. 댓글도 많이 달았는데~~~ 2015에는 시급 좀 많이 올려주세요. ㅋㅋㅋ
아... 라이모도 시급 좀 올려 달래요. 하루옹이랑 비머는 시급 안올려줘서 삐친 듯 함Yo~~
지난주에도 여럿. . 학대(?)광경을 너무 확대해서 보믄 손해~!^^
(우쒸... 실험 사진이 안올라가네... 새글 추가~)
하루랑 비머랑 친해진거~? 혹시~? ㅋㅋㅋㅋㅋ 이 잉간덜 김씨랑 똑같이 전화도 안받는 넘들 되었다는~~~
@피노이(박천형) ㅋㅋㅋ 시급이 넘 짜요~~ "짜요~~"
쥐가 불쌍해요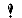
@.@;;
헉
학문에 대한 열정이 대단하십니다. 저는 아직 젊은 나이지만 논문은 엄두를 못내고 있습니다.